Микроорганизмы, токсины и эпидемии. 3.3. После СПИДа
Дальнейшее развитие пандемии СПИДа порождает две взаимоисключающие пандемические тенденции:
1. ВИЧ является только одним из представителей лентиовирусов, вызвавших в XX веке обширные эпидемические процессы в популяциях людей и домашних животных. Между представителями данного семейства вирусов также возможна конкуренция за территорию своего обитания, в результате которой в их экологической нише останется лишь наиболее приспособленный вид (в эволюционной биологии этот процесс называется поиском оптимального фенотипа). Кроме того, конкуренция возможна между ретровирусами и другими мобильными «обитателями» хромосом приматов, а также между другими микроорганизмами (отдельные ДНК-вирусы, микоплазмы и др.), использующими вторую стратегию паразитизма.
2. Распространение ВИЧ/СПИДа, лейкозов, сывороточных гепатитов, широкое использование в клинической практике цитостатиков и иммунодепрессантов в совокупности ведут к постепенному снижению иммунной защиты у миллионов компактно проживающих представителей вида Homo sapiens и создают условия для проникновения, а потом и его «заселения» новыми микроорганизмами-паразитами. Кроме того, в этих условиях значительно облегчится отбор и формирование высокоспециализированных эпидемических вариантов среди микроорганизмов, ранее таковыми не считавшимися. Поэтому, при достижении иммунодефицитными популяциями определенного порогового уровня или слияния их в одну, достаточно крупную (глобальную?) популяцию, весьма вероятно возникновение эпидемий, вызываемых «побежденными» и «контролируемыми» микроорганизмами, использующими первую стратегию паразитизма, а также их близкородственными видами и совершенно неожиданными для здравоохранения развитых стран возбудителями.
Развитию обеих тенденций способствует широкое использование в XX веке для борьбы с инфекционными болезнями средств иммунопрофилактики и антибиотико(химио)терапии. Способствуя (в краткосрочной перспективе) победам над отдельными инфекционными болезнями, одновременно они создают условия для эволюционного отбора видов, паразитирующих в новых условиях. Возможности технологий, используемых для дальнейшего совершенствования таких средств не безграничны, и неизбежно будут исчерпаны. В результате станет невозможным создание лечебных и профилактических препаратов, позволяющих сдерживать развитие запущенных ими же эпидемических процессов.
Возбудитель, способный «заменить» ВИЧ. Внутренняя логика и преемственность приведенных в эволюционной модели пандемий событий (см. рис. 64), позволяют предположить, что следующая пандемия («после СПИДа») будет вызвана микроорганизмом, использующим третью стратегию паразитизма. Такой паразит не только внедряется в геном хозяина и становится генетическим паразитом, но и активно разрушает его участки или отдельные гены, важные для жизнедеятельности человека (табл. 32).
Таблица 32
Возможные свойства паразита, способного вытеснить ВИЧ*
|
Характер инфекционного процесса (по сравнению с ВИЧ) |
Решение определенных эволюционных задач |
Социальные факторы, способствующие появлению возбудителя |
|
Кроме полового и гематогенного, основным механизмом передачи паразита станет наследственный |
Паразит обретает «бессмертие» и будет существовать столько же, сколько вид Н. sapiens. При этом значительно снижается риск его «не передачи» половым или гематогенным путем либо гибели в результате внезапного вымирания всей иммунодефицитной популяции из-за какой-то быстрой контагиозной инфекции |
Профилактические мероприятия, связанные с ранним выявлением СПИДа; выбраковка партий крови, содержащих ВИЧ; ограничение возможностей инфицирования при инъекциях и половых контактах |
|
Увеличение возможности паразита по интеграции в различные участки генома человека, а также увеличение числа сайтов интеграции |
Вытеснение ВИЧ из его «экологической ниши», расширение своей собственной |
Использование технологии генотерапии для лечения и профилактики СПИДа, плюс те же факторы, которые способствуют проникновению в человеческую популяцию ВИЧ |
|
Кроме присоединения (или вместо нее) вторичной инфекции, разрушение отдельных генов, нарушение регуляции метаболических процессов, неопластические изменения и другая патология, связанная с вмешательством в работу генома человека |
Сохранение способности к персистированию паразита в условиях противодействия репарационной и других подобных систем хозяина, рассматривающих его как вредную мутацию (блокирование репарационных механизмов макроорганизма, конкуренция с генами — трансактиваторами ВИЧ, перемещение по геному с захватом участков ДНК, интеграция в гомеозисные гены, решение задачи собственного «бессмертия» на уровне клеток отдельной ткани и др.) |
Сохранение генетически дефектных (инфицированных) особей, максимальное продление сроков их жизни с возможностью репродукции, отсутствие контроля со стороны общества за размножением их потомства |
* Supotnitskii, 1995
Способность вирусов разрушать гены при интеграции в геномы эукариот, явление известное [Филдс Б. и др., 1989]. Однако в данной ситуации этот процесс будет следствием конкурентной борьбы ретровирусов за влияние над геномом, как между собой, так и с теми мобильными элементами хромосом, которые, возможно, играют роль «защитных экранов» генома от таких паразитов. Преимуществом будет обладать паразит, способный к более эффективной интеграции и противодействию защитным системам хозяина. Увеличение же числа сайтов интеграции, блокирование репарационных систем макроорганизма, конкуренция с генами-трансактиваторами (трансрегуляторами) ВИЧ, перемещение по геному с захватом участков ДНК, интеграция в гомеозисные гены (регуляторные гены высшего порядка, управляющие работой других генов), приведут к возникновению патологии, которая далеко не сразу будет воспринята учеными как инфекционная.
Учитывая то, что ВИЧ интегрируется с геномом человека, можно предположить, что именно в рамках технологий, используемых для генотерапии, удастся разработать эффективные подходы к лечению и профилактике СПИДа. Но когда какие-то участки генома человека, являющиеся сайтами для интеграции ВИЧ, либо геном самого вируса станут мишенями для генотерапевтов, возникнут условия для прямого селективного давления на популяции ретровирусов. В свою очередь сами ретровирусы не являются однородными и содержат различные варианты даже одних и тех же возбудителей. Из этих вариантов, «выдавленных» из традиционных для них участков генома, может начаться формирование патогенов, интегрирующихся с участками генома, которые какое-то время будут недоступны для генотерапевтов, например, в гомеозисные гены. Нарушение функции последних равноценно разрушению больших участков генома. Возможно появление ретровирусов, не имеющих определенных сайтов интеграции или (наихудший для данной ситуации вариант), не проявившихся в первом поколении их новых хозяев.
Из существующих возбудителей инфекционных болезней такую пандемию может вызвать малоизвестная (или неизвестная еще) разновидность онковирусов, принадлежащая к семейству ретровирусов и попавшая каким-то образом в человеческую популяцию из животного резервуара (например, в результате опытов по трансплантации органов и тканей; использования вакцин, приготовленных из культур клеток животных и др.), либо образовавшаяся в результате рекомбинации вирусов, освоивших одну экологическую нишу (например, имеющих общие сайты интеграции в геноме) и по выше указанным причинам, получившая селективные преимущества перед ВИЧ.
Что касается особенностей новой пандемической ситуации, то хотелось бы обратить внимание на тот факт, что и интеграция ВИЧ с геном человека не проходит для последнего бесследно. Уже в первом описании заболевания СПИДом отмечена большая смертность больных от опухолей, в частности, от рака эндотелия сосудов, что в какой-то мере служит косвенным подтверждением избирательного поражения ВИЧ отдельных генов человека. Имеются прямые экспериментальные подтверждения этого факта. Показано, что введение мышам регуляторного гена tat ВИЧ вызывает у них патологическое состояние, напоминающее саркому Капоши [Cichutek К., 1992]. На основании вышеизложенного можно сделать предположение, что процесс формирования подобного возбудителя уже начался в подсемействе лентиовирусов, хотя у болезни, вызываемой наиболее распространенным в настоящее время их представителем, совершенно иной патогенез.
Клинические проявления новой болезни представить довольно трудно. Ими могут быть быстро прогрессирующие неоплазии, иммунодефициты, мышечная дистрофия, демиелинизация, отложения бета-амилоидных белков, психические нарушения, дефекты развития и другие поражения генетического характера. Если исходить из того, что новый возбудитель будет способен осваивать экологические ниши вытесняемого им ВИЧ, то наиболее вероятна патология, связанная с дефектами ДНК Т-лимфоцитов, эндотелиоцитов кровеносных и лимфатических сосудов, эпителиальных клеток кожи, астроцитов и нейронов мозга.
Инфекционистам клинические проявления болезни могут напоминать СПИД. Но им не удастся подтвердить диагноз фактом обнаружения антител к ВИЧ-1 или ВИЧ-2 и добиться улучшения состояния больного обычно применяемым азидотимидином. Инкубационный период, по-видимому, будет превышать таковой у ВИЧ/СПИДа. К механизмам переноса нового возбудителя может добавиться еще один — наследственный.
Такой путь передачи вирусов не является для высших животных чем-то новым. Предполагается, что генетическим путем от матери к потомству через эмбрион передается вирус лейкоза мышей Гросса и Биттнеровский агент карциномы молочных желез [Букринская А. Г., Жданов В. М., 1991]. Отсюда возможная длительность пандемического процесса — от нескольких десятилетий и, вполне вероятно, до столетий, а при активном использовании технологий генотерапии и формировании трансгенных популяций людей — частичное стирание различий между патологией наследственной и инфекционной. Сформируется новая модель пандемического процесса, когда возбудитель болезни будет способен передаваться между людьми, не попадая в окружающую среду. Отдаленным последствием пандемического распространения такого паразита станет формирование обширных популяций генетически обреченных людей.
Приведенные выше данные одновременно свидетельствуют и о возможности существования механизма случайного (т. е. не обусловленного резким ухудшением условий обитания) самоуничтожения любого эукариотического вида, действующего на генетическом уровне.
Метка Господа. Еще в XIX столетии известный русский богослов А. П. Лопухин (1887) заметил, что в древнем Египте предание о потопе изменено было в том смысле, что «нечестивый род человеческий наказан был язвой, истребившей его весь, за исключением немногих лиц, сделавшихся родоначальниками нового человечества». Наличие водных путей по притокам Нила из района современной Уганды, эндемичного как по ВИЧ, так и по опасным для людей осповирусам, позволяют отнестись к этому древнему мифу серьезно. СПИД имеет все признаки давно сформировавшейся болезни. Учитывая способность ВИЧ распространяться при невысокой плотности населения, можно предположить, что в первобытный период нашей истории инфицирование им людей было значительно большим, чем сейчас, и ВИЧ был мощным, длительно действующим фактором, сдерживавшим размножение вида Homo sapiens (аналогичное явление сегодня обнаруживают в стадах некоторых приматов).
Однако ВИЧ открывал ворота для проникновения в популяции вида Homo sapiens возбудителей, называемых сегодня «оппортунистическими». Среди таких микроорганизмов могли оказаться неспециализированные предковые формы многих из тех, кого сейчас мы считаем строгими антропонозами, т. е. вызывающими болезнь только у людей. Объясним это на примере возможного формирования возбудителя натуральной оспы.
Задолго до появления вируса натуральной оспы, в результате эволюционной дивергенции в семействе Poxviridae произошло обособление отдельных видов в разных экологических нишах. Это снимало конкуренцию между подвидами и вело к образованию неконкурирующих между собой видов, при относительно небольшой генотипической дифференциации между ними. Малочисленность и разбросанность популяций диких животных позволяли распространяться среди них возбудителям, вызывающим оспенные инфекции с длительным заразным периодом, наличием носительства и хроническим течением болезни. Проникнув случайным образом в популяцию вида Homo sapiens, микроорганизм либо не вызывал специфической болезни, либо не мог серьезным образом конкурировать в эпидемических цепочках с другими патогенными микроорганизмами человека. Иная ситуация могла наблюдаться в достаточно крупных иммунодефицитных сообществах людей (рис. 66).
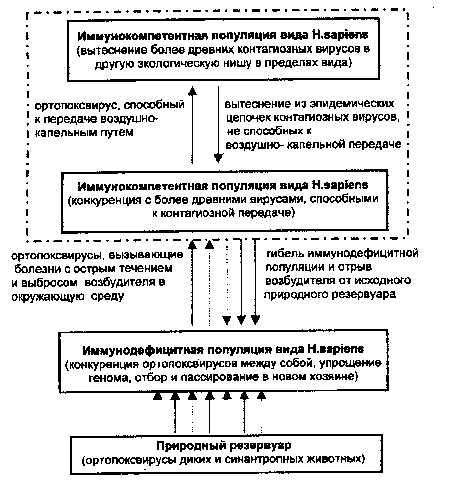
Рис. 66. Схематическое изображение возможного механизма формирования узкой специализации у возбудителя натуральной оспы и его отрыва от исходного природного резервуара.
G. Werner и соавт. (1968), еще до обнаружения ВИЧ, провели ряд интересных экспериментов, показавших, что у иммунодефицитных особей возможно значительное снижение специфичности заражения поксвирусами. По их данным у облученных кроликов вирус вакцины (ортопоксвирус, используемый для иммунизации против вируса натуральной оспы) вызывал острую болезнь. У облученных крыс, обычно абсолютно невосприимчивых к вирусу этромелии (вирус оспы мышей), инфицирование этим вирусом вызывало острую инфекцию с 30% смертностью. Аналогичное явление могло наблюдаться и среди иммунодефицитных людей, только с той поправкой, что инфекционные процессы происходили при жесткой конкуренции между несколькими видами ортопоксвирусов, чьи экологические потребности оказались сходными. Полиморфизации вирусов, накоплению их специализированных дегенеративных форм, способствовало отсутствие селективного давления со стороны иммунной системы. В эпидемических цепочках, образовавшихся в социально организованных и плотных «популяциях» людей, большую возможность сохраниться имели подтипы возбудителей, вызывающих болезнь с острым течением и массивным выделением вируса в окружающую среду. Одновременно, пассированием через особи с различной степенью иммунодефицитности, происходил отбор более вирулентных производных. В жесткую конкуренцию с поксвирусами могли вступить более древние возбудители эктодермозов, например, из семейства Herpesviridae, также способные к контактной передаче. В этой борьбе победил вирус, передающийся между людьми воздушно-капельным путем и имеющий относительно длительный инкубационный период. Последнее свойство позволило ему, инфицируя подрастающее восприимчивое поколение, сохраняться тысячи лет в относительно малочисленных и приобретших постэпидемический иммунитет популяциях.
Образование в процессе эволюции вируса натуральной оспы из циркулирующих в популяциях диких животных менее специализированных осповирусов, разумеется, было случайным событием. Однако он оказался идеальным антагонистом возбудителей медленных инфекций, вызывающих развитие иммунодефицитных состояний. Инкубационный период оспы достигает 17 сут и поэтому болезнь распространяется медленно, но благодаря высокой контагиозности возбудителя — по принципу «из дома в дом», поражает многих.
В отличие от ВИЧ, вирус натуральной оспы менее опасен, он не вызывает 100% гибели заболевших людей и не образует столь длинных (ограниченных лишь количеством человеческих особей) эпидемических цепочек. Постепенное и повсеместное истребление ВНО носителей ретровирусов среди людей, оборвало их эпидемические цепочки. Это способствовало увеличению продолжительности жизни и репродуктивного периода человека. Отсюда, видимо, идет и происхождение древнеегипетского мифа, произвольно истолкованного Лопухиным как свидетельство о потопе; и представление об оспе у всех народов, сталкивавшихся с ней, как об «очистительной болезни»; а также почитание Бога Оспы отдельными племенами, расселение которых в Центральной Африке происходило в античное время и позже (рис. 67).

Рис. 67. Бог оспы Сопоне. Бог оспы Сопоне — один из наиболее почитаемых богов у нигерийского племени йоруба [Девис М., 1980]
На протяжении истории человечества оспа появлялась и исчезала неоднократно (см. рис. 21). По завершению последней великой оспенной пандемии в конце XVII столетия, во всем мире трудно было найти взрослого человека без оспенных отметин на лице. В те годы оспины рассматривались как в своем роде «метка Господа», даровавшего этому человеку жизнь. Английский историк Маколей писал про те времена: «Моровая язва или чума была более смертельна, чем оспа, но зато она посещала наши берега лишь однажды или дважды на памяти людей, тогда как оспа неотступно пребывала между нами, наполняя кладбища покойниками, терзая постоянно страхом всех тех, которые еще не болели ею, оставляя на лицах людей, жизнь которых она пощадила, безобразные знаки, как клеймо своего могущества» (цит. по К.В. Рыжову, 1999). Начало же последнего периода экспоненциального роста численности человечества, многие ученые хронологически относят на конец XVII столетия, т. е. на окончание последней великой оспенной пандемии.
Генетические метки прошлых пандемий СПИДа. Выше мы уже отметили, что повышенную частоту встречаемости 32 bр делеций гена CCR5 в популяциях людей, можно рассматривать как своеобразный генетический шрам, оставленный прошлыми эпидемиями СПИДа. Но африканцы, контактирующие с природным резервуаром ВИЧ, находящимся среди африканских приматов, оказывается, менее знакомы со СПИДом, чем европейцы! Это заставляет усомниться в правильности распространенных представлений о месте происхождения негроидной расы. Частота распределения аллелей гена CCR5 свидетельствует о том, что люди черной расы пришли в тропическую Африку сравнительно недавно и с севера, предки же современных европейцев контактировали с ВИЧ очень длительное время. Однако где и когда это произошло?
В какой-то мере ответ на этот вопрос дает отсутствие следов цивилизаций в регионе Африки южнее Сахары. Считается, что центральная и восточная Африка являются очагами зарождения человечества, однако каких либо следов материальной культуры, появившихся там ранее I в. н. э., южнее Сахары не находят [см. работу Бонгард-Левиной Г.М., 1989]. В европейских популяциях людей ВИЧ присутствовал еще с тех времен, когда разрозненные и малочисленные праевропейские племена шли за отступающими ледниками на север Европы. Из-за небольшой численности и рассеянности они были менее уязвимы для возбудителей острых контагиозных инфекций (т. е. использующих стратегию паразитизма первого типа), в том числе и ВНО. Однако эпидемические цепочки ВИЧ и подобных вирусов среди европейцев поддерживались сравнительно долго, возможно до античных и, даже до сокрушительных раннесредневековых эпидемий контагиозных болезней, собирательно называемых тогда «чумой». До эпидемий контагиозных болезней продолжительность жизни «первобытных» европейцев редко превышала 30 лет, как сегодня негров в пораженной ВИЧ Уганде.
Формирование первых государств в истории человеческой цивилизации (V—IV тысячелетие до н. э.), происходило именно вдоль путей естественного распространения ВНО из регионов Центральной Африки (долина Нила, Ближний Восток, Месопотамия). Люди негроидной расы, пришедшие в Центральную Африку с севера уже после образования там ВНО, подвергались его селективному давлению на протяжении нескольких тысяч лет. И это давление продолжалось до проведения полной противооспенной иммунизации населения региона. Эпидемические цепочки, тянущиеся к людям от резервуаров ВИЧ в популяциях приматов, весь этот период были блокированы ВНО. Поэтому африканцы не имеют мутантных аллелей гена CCR5 и более чувствительны к ВИЧ, чем европейцы.
Глобальный пандемический цикл. На рис. 68 представлена модель глобального многовекового цикла быстрых и медленных инфекций.
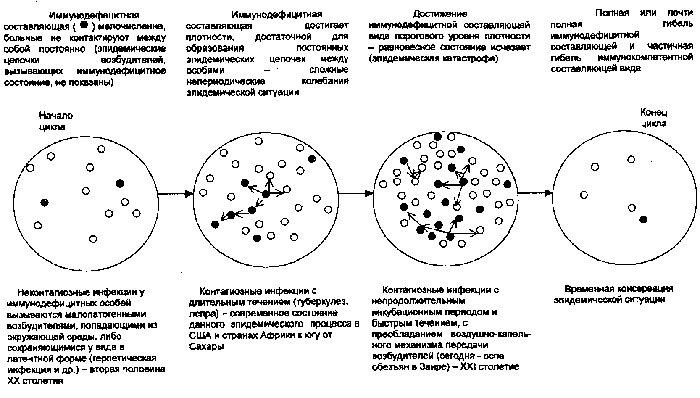
Рис. 68. Схема глобального многовекового цикла быстрых и медленных инфекций
По мере увеличения численности иммунодефицитных людей в популяции, наибольшие преимущества в эпидемических цепочках получают возбудители контагиозных инфекций. Цикл заканчивается масштабными, трудно контролируемыми эпидемиями контагиозных инфекций, вызываемыми возбудителями с коротким инкубационным периодом (в нашем восприятии времени) и массированным выделением возбудителя в окружающую среду. Такие эпидемии могут охватывать отдельные регионы и страны, вызывать огромные опустошения и тянуться, периодически затихая, десятилетиями. Затем они прекращаются и численность населения быстро растет.
Глобальное распространение туберкулеза является проявлением такого цикла для ситуации, когда плотность иммунодефицитной составляющей достаточна для образования устойчивых эпидемических цепочек контагиозных инфекций с длительным течением (туберкулез, проказа), но еще не достигла «критической массы» необходимой для появления контагиозных инфекций с коротким инкубационным периодом (в нашем восприятии времени). Однако такая ситуация уже складывается на севере Заира, где плотность ВИЧ-инфицированного населения достигает 10%. Эндемичный для этих мест возбудитель оспы обезьян начал передаваться не только от обезьян к людям, но и между людьми (рис. 69).
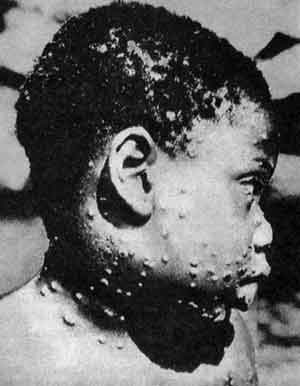
Рис. 69. Оспа обезьян у жителя Заира. Обнаруженный еще в 1970-м г. в Заире мальчик, заболевший оспой обезьян. Тогда, на фоне побед над ВНО, эти случаи казались досадным недоразумением. Считалось, что оспа обезьян малоконтагиозна и большой опасности не представляет. Однако позже, вопреки всем своим прежним заявлениям о прекращении глобальной иммунизации против оспы, ВОЗ возобновила иммунизацию населения тропических регионов Африки, и не афишируя продолжала ее до средины 1990-х гг., когда из-за резко возросшего числа ВИЧ-инфицированных людей, иммунизация стала невозможна [Cohen J., 1997]. Т. е. ситуация по оспе, теперь уже не важно по какой — оспы обезьян или натуральной, в регионах Африки, эндемичных по ВИЧ, постепенно становится неуправляемой.
По мере совершенствования средств борьбы с возбудителями инфекционных болезней, цикл становится все более растянутым, но последствия пандемий все более губительными.
В небольшой заирской деревушке, в районе Котако-Комба, оспой обезьян заболело 42 жителя из 346, трое из них погибло. Установлен больной, ставший источником каскада передач вируса 8 человекам, что уже сопоставимо с контагиозностью ВНО. Все заболевшие ранее не были иммунизированы против ВНО [Mukinda V. et al., 1997].
***
В отсутствие селективного давления со стороны иммунной системы человека происходит полиморфизация популяции ВИЧ и ВИЧ-подобных вирусов, проникновение других ретровирусов, что ведет к конкуренции между ними, их отдельными подтипами, эндогенными ретровирусами и другими мобильными элементами человеческого генома. Конечной задачей такой конкуренции ретровирусов и ретроэлементов между собой является усиление влияния на геном хозяина. Поэтому можно предполагать, что дальнейшее развитие пандемии СПИДа приведет к появлению новой инфекционной патологии, связанной с избирательным поражением жизненно важных участков генома человека.
При достижении иммунодефицитной составляющей человеческого общества пока еще не известной «критической массы», ситуация может неожиданно разрешиться глобальной эпидемической катастрофой, по типу наблюдавшихся в средние века (к сожалению, для человечества это более благоприятный вариант развития событий).
Обе тенденции проявляют себя тем, что идет малозаметный процесс формирования возбудителей, обладающих новым патогенным потенциалом. Масштабное проникновение новых и возвращение «старых» возбудителей инфекционных болезней в человеческое общество станет возможным тогда, когда будут исчерпаны возможности технологий, используемых для создания новых средств иммунопрофилактики, антибиотико- и химиотерапии. Видимо этот механизм является естественным регулятором пандемий, вызываемых ВИЧ и другими микроорганизмами, использующими вторую и третью стратегии паразитизма.