Микроорганизмы, токсины и эпидемии. 3.2. СПИД
Распространение среди людей «неконтагиозного» ВИЧ — возбудителя СПИДа, произошло на фоне успехов в борьбе с инфекционными болезнями и, причем столь стремительно, что невольно приходишь к выводу о существовании очень серьезных оснований для этого процесса.
Появление возбудителя СПИДа. На основе анализа нуклеотидных последовательностей ретровирусов, выдвинуто несколько взаимоисключающих гипотез о возможных сроках формирования вируса, возбудителя СПИДа. Данные, полученные В. Mager (1987), позволили ему предположить, что ВИЧ, как патогенный для человека паразитический микроорганизм, сформировался в Африке сравнительно недавно — в 1950-е гг. D. Xuminer и соавт. (1987) сделали ретроспективный анализ медицинской литературы и выявили 14 случаев инфекционных болезней, описанных до 1981 г, которые соответствовали критериям, предложенным для СПИДа. Но география выявленных ими случаев болезни не ограничивалась Центральной Африкой, а включала США, Канаду, Великобританию, Израиль, Бельгию и некоторые другие страны. Наиболее же «старый» случай наблюдался в 1953 г. в США [Xuminer D. et al., 1987]. Его основным проявлением была цитомегаловирусная пневмония, обычно возникающая только на девятый год течения болезни [Милдз Дж., Мазур Г., 1990]. Следовательно, инфицирование людей ВИЧ было возможным уже в середине 1940-х гг. — во время миграционных волн Второй мировой войны, что противоречит датировке появления эпидемического варианта ВИЧ, сделанной В. Mager (1987). Отсюда можно сделать еще один вывод — если ВИЧ и начал мутировать только в 50-е годы ХХ столетия, то это не объясняет малую эпидемическую значимость вируса в предшествующие годы. Слишком простым выглядит объяснение возникновения пандемии СПИДа перемещением в африканские города, фабричные поселки и рудники избыточного мужского населения из деревень, предложенное N.M. Ampel (1991). По своей сути оно подразумевает те же механизмы распространения инфекционной болезни, которые наблюдаются при вспышках чумы и туляремии в годы высокой численности грызунов. В течение нескольких столетий из Африки были вывезены миллионы рабов. Приобрети тогда среди них СПИД характер массовой болезни, он был бы неизбежно описан в виде отдельных патологических синдромов, снижающих качество «товара».
Более вероятно длительное существование какого-то мощного фактора, сдерживавшего широкомасштабное распространение ВИЧ из эндемических регионов. Устранение этого фактора привело к формированию столь крупных популяций ВИЧ-инфицированного населения в неэндемичных по данному вирусу регионах мира, что возбудитель потерял связь со своим первичным резервуаром. Однако обратимся к данным других исследователей.
Высокий уровень инфицированности населения к югу от Сахары позволил Р. Андерсону и Р. Мейю (1992) предположить, что ВИЧ там существует уже давно. В отличие от В. Mager (1987), по результатам анализа молекулярной структуры вируса они определяют этот срок в 100—200 лет и более. Он согласуется с наблюдениями D. Xuminer и соавт. (1987). Но тогда, что же способствовало ускорению мутирования и распространения ВИЧ в 60-е и 70-е гг. ХХ столетия?
Ответ на этот вопрос мы видим в следующем явлении. Начало обоих процессов по времени отчетливо совпадает с резким снижением заболеваемости населения региона натуральной оспой. В отличие от Северной Африки, Европы и Американского континента, оспа и СПИД в странах к югу от Сахары были не заносными, а являлись эндемическими болезнями [Андерсон P.M., Мей P.M., 1992; Шувалова Е.П., 1973]. И население именно тех стран, где сейчас повсеместно распространен ВИЧ, к началу 1960-х гг. было поражено вирусом натуральной оспы в значительно большей степени, чем в других регионах Африки [Бароян О.В., 1967; Шувалова Е.П., 1973]. В этих условиях люди, больные СПИДом, не могли долго существовать. Эндемический вирус натуральной оспы обрывал цепочки, по которым распространялся эндемический для данного региона ВИЧ. В результате ВИЧ не имел возможности эволюционировать, его законсервированные в более древних гоминидах варианты ничем не успевали проявляться у людей на фоне других массовых инфекций (рис. 65).
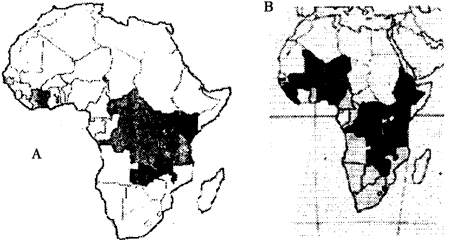
Рис. 65. Совпадение ареалов ВНО и ВИЧ в центральных районах Африки. Ареал, где сегодня особенно сильно распространен СПИД (карта А; Андерсон Р., Мей. Р., 1992), аккуратно располагается на территориях, ранее «занятых» натуральной оспой (карта В; Eradication de la variole..., 1968). Участники Всемирной ассамблеи здравоохранения, которая вынесла в 1966 г. решение об интенсификации программы ликвидации натуральной оспы во всем мире, не могли себе даже представить, что через 20 лет именно в тех регионах Африки, где у них был наибольший успех, образуется самая крупная иммунодефицитная «популяция» вида Homo sapiens. Co средины 1990-х гг. в ареале СПИДа на севере Заира отмечено возрастание количества людей, инфицированных вирусом оспы обезьян и повышение контагиозности этого вируса до показателей ВНО. При этом генетически вирус оспы обезьян за 20 лет не изменился. Повысилась восприимчивость к нему людей, проживающих в регионе [Cohen J., 1997].
Противооспенная иммунизация, длительно проводимая скарификационным методом (т. е. нанесением вакцины на царапины!) в районах, для которых ВИЧ и вирус натуральной оспы были эндемичны, и последовавшее за этим сокращение заболеваемости населения натуральной оспой, одновременно способствовали продлению жизни людей — больных СПИДом.
Отсутствие же селективного давления на ВИЧ, а также подавление в развитых странах с помощью вакцинопрофилактики и антибиотико(химио)терапии других возможных конкурентов ВИЧ — возбудителей быстрых инфекций (использующих первую стратегию паразитизма), благоприятствовали распространению в человеческих популяциях штаммов вируса, вызвавших в конце XX столетия пандемию СПИДа.
А теперь посмотрим, кого же мы все-таки победили в 1970-х гг.? Наиболее тяжелые эпидемии натуральной оспы отмечены в XVII и XVIII веках. Тогда наличие иммунного населения, т. е. тех, кто переболел оспой, не сдерживало развитие новых эпидемий. По истечении определенного срока подрастало новое восприимчивое к оспе поколение. Люди старшего возраста постепенно утрачивали иммунитет, приобретенный ими в детстве, и циркуляция вируса не прекращалась. Результативность и безопасность вакцины Дженнера оспаривались не только его современниками. На протяжении всего XIX столетия находились ученые, аргументировано отрицавшие целесообразность массовой противооспенной вакцинации. Но так как с начала XIX столетия смертность в очагах натуральной оспы постепенно шла на убыль, доверие к вакцине возрастало. К концу XIX столетия в ее эффективности уже почти (!) никто не сомневался.
Ликвидация натуральной оспы в 1960-1970-х гг. в странах, находящихся к югу и юго-востоку от Сахары, происходила на фоне широко распространившихся менее опасных штаммов вируса (типа minor). Вызываемые такими штаммами оспоподобные болезни под разными названиями (аластрим, санага, самоа и др.) были известны ранее, однако длительное время они не были преобладающими. Выполненные несколько лет назад молекулярно-генетические исследования вирусов, выделенных в последних очагах натуральной оспы, показали, что геномы менее вирулентных для людей штаммов отличаются от геномов высоковирулентных возбудителей натуральной оспы увеличением размеров инвертированных терминальных повторов (ITR). В тоже время основная часть генома вируса остается неизмененной (табл. 31).
Таблица 31
Характеристика вирусов, выделенных в последних очагах натуральной оспы
|
Изолят (страна), год выделения |
Тип |
Смертельные исходы, % |
Размер ITR, пары оснований |
|
Harvey (Англия), 1944 |
major |
27,3 |
518 |
|
Bangladesh (Бангладеш), 1975 |
major |
18,5 |
725 |
|
Congo (Конго), 1970 |
major |
9,6 |
793 |
|
Somalia (Сомали), 1977 |
minor |
0,4 |
1051 |
R. Massung и соавт., 1995.
Эти данные свидетельствуют о каком-то самостоятельном процессе — приспособительном мутировании самого вируса в направлении образования маловирулентных штаммов, не проявляющих себя вспышками болезни, сопровождающихся массовой гибелью людей. Считающийся последним очаг оспы, ликвидированный в 1977 г. в Сомали, был вызван штаммом типа minor. Смертность в очаге не превышала 0,4 %. И если быть строгим в терминах, то эта болезнь уже не была натуральной оспой, а представляла собой оспоподобное заболевание — аластрим. Следовательно, возбудитель натуральной оспы был «побежден» тогда, когда он, как это уже не раз было в прошлом, по каким-то непонятным для нас причинам уходил сам.
Заметим, что аналогичное явление — «самопроизвольное» снижение вирулентности и контагиозности, и «уход» из человеческих популяций, подмечено исследователями для многих других, весьма смертоносных в прошлом возбудителей инфекционных болезней (сифилиса, проказы и др.). Но против них никогда не осуществлялись программы глобальной иммунизации.
Так для кого же освободил место вирус натуральной оспы в глобальном масштабе?
Особенности пандемии ВИЧ.
Распространение ВИЧ не требует высокой плотности населения, поддерживающей непрерывность пандемий, вызываемых возбудителями оспы или гриппа. Растянутость пандемического процесса по времени (в нашем его ощущении) в данном случае не формирует устойчивой популяции, а увеличивает ее инфицированность. Длительность носительства (опять же в нашем ощущении времени) и отсутствие стойкого постинфекционного иммунитета, компенсируют низкую инфекционность самого вируса и отсутствие его массивного выделения в окружающую среду с заражением большого числа людей. Инфицированный человек становится заразным для других людей довольно быстро. По некоторым данным, переливание крови от зараженного лица уже через 1–2 недели после предполагаемого заражения, приводит к развитию инфекционного процесса у реципиента. Заболевшие не изолируются и вследствие развившейся деменции могут не воспринимать критично свое состояние, оставаясь источником возбудителя до конца жизни. Неинфицированная часть населения из-за законодательных ограничений не может избегать контактов с такими больными и полностью зависит от их доброй воли. Дети, родившиеся от ВИЧ-инфицированных матерей, как правило, тоже инфицированы либо инфицируются позже, во время ухода и кормления. Эффективные карантинные мероприятия при этой болезни невозможны из-за ее длительного инкубационного периода (в нашем ощущении времени). Контролировать распространение инфекции средствами иммунопрофилактики вряд ли когда удастся. Вирус существует в организме больного в двух формах — РНК-содержащей и ДНК-содержащей. Последняя встроена в геном хозяина и не имеет антигенов, доступных для иммуноглобулинов. Кроме того, работы по созданию средств и методов лечения и профилактики болезни наталкиваются на отсутствие соответствующих экспериментальных моделей. Увеличению инфицированности населения способствует и паллиативный характер лечения больных СПИДом. Продление их жизни без полного излечения, а только за счет удлинения инкубационного и симптоматического периодов болезни увеличивает резервуар возбудителя и ведет к тому, что скорость его распространения все более опережает смертность от него. Как и в средние века: «Ничего не могли с ней поделать догадливость и предусмотрительность человеческая...».
Неизвестные виды ВИЧ.
На 8-й Международной конференции по проблеме СПИДа (1992) наибольшее внимание привлекло сообщение о пациентах с прогрессирующей супрессией иммунной системы при отсутствии у них ВИЧ-1 и ВИЧ-2. Полагают, что новый вирус передается легче, чем вирус, вызывающий СПИД [Baum R., 1992]. Любопытно и то, что сообщения о так называемой идиопатической СД 4 лимфопении («ВИЧ отрицательный СПИД») в последующие годы появились в США [Ramirer J. A. et al., 1994], в Германии [Neukirch В., Kremer G. J., 1995], в Испании [Sancher R. et al., 1994]. Распространение этого, пока необъяснимого заболевания, все больше приобретает параллельный «ВИЧ положительному СПИДу» характер. Так же есть сообщение о том, что причина аутоиммунной болезни человека, называемой синдромом Шегрена, является инфицирование ретровирусом, антигенно сходным с ВИЧ [Carri R. et al., 1992].
Патогенные ретровирусы распространяются в последние годы и среди домашних животных. Глобальное значение приобретает инфекционное заболевание висна-меди, поражающее овец. У вируса висны и ВИЧ общий предок [Букринская А. Г., Жданов В. М., 1991]. До 7% крупного рогатого скота в развитых странах уже поражено бычьим вирусом иммунодефицита [Федоров Ю. Н., Верховский О. А., 1996].
***
Характер вызываемых ретровирусами эпидемических процессов как нельзя лучше соответствует направленности селективного отбора паразитических видов в условиях жесткого давления на микроорганизмы, использующие первую стратегию паразитирования. В отличие от других эндогенных транспозиционных элементов генома человека (например, ретротранспозонов), ретровирусы способны к цитоплазматической репликации. Поэтому они размножаются быстрее, чем популяция хозяина, и, следовательно, при наличии механизма передачи способны заразить ее всю на протяжении смены какого-то количества поколений. Продолжительность этого процесса с эволюционной точки зрения никакого значения не имеет. Человечество уже неоднократно встречалось с ВИЧ и подобными вирусами, однако, вызванные ими эпидемии самоограничивались существующими в природе механизмами.