«Черная смерть» — механизм пандемической катастрофы

Предназначалась для журнала «РЭТ-инфо». В состав этой статьи входит ПРИЛОЖЕНИЕ 1 - содержащее мнение членов Парижского медицинского факультета о происхождении эпидемии чумы черной смерти и о предохранительных мерах против нее.
«Братья мои! – возгласил он с силой. - Эта смертоносная охота идет ныне на наших улицах. Смотрите, смотрите, вот он, ангел чумы, прекрасный, как Люцифер, и сверкающий, как само зло, вот он, грозно встающий, над вашими кровлями, вот заносит десницу с окровавленным копьем над главою своею, а левой рукой указует на домы ваши. Быть может, как раз сейчас он простер перст к вашей двери, и копье с треском вонзается в дерево, и еще через миг чума входит к вам, усаживается в комнате вашей и ждет вашего возвращения. Она там, терпеливая и зоркая, неотвратимая, как сам порядок мироздания. И руку, что она протянет к вам, вам ни одна сила земная, ни даже – запомните это хорошенько! – суетные человеческие знания не отведут от вас. И поверженные на обагренное кровью гумно страданий, вы будете отброшены вместе с плевелами».
Из проповеди отца Панлю (Альбер Камю «Чума», 1947)
«От чего они умерли? – Спендер подошел ближе.
Вы не поверите.
Что их убило?
Ветрянка, - коротко ответил Хэтуэй.
Не может быть!
Точно. Я сделал анализы. Ветряная оспа. Ее действие на марсиан совсем не такое, как на землян. Видимо, все дело в обмене веществ. Они почернели, как головешки, и высохли, превратившись в ломкие хлопья. Но это ветрянка, никого сомнения…»
Рей Бредбери «Марсианские хроники» (1950)
Сокрушительная пандемия чумы 1346-1351 гг., названная современниками черной смертью , до сих пор не нашла удовлетворительного объяснения. Попытки понять ее причины обычно сводятся либо к поиску доказательств того, что это была не чума (Lederberg J., 1997), и охотно принимаются учеными, озабоченными проблемой появления новых инфекций. Либо из контекста исторических событий извлекаются те исторические источники, которые можно интерпретировать как свидетельство применения биологического оружия (БО) татарским ханом Джаныбеком (Kortepeter M. et al., 2001; Wheelis M., 2002 и др.). Разумеется, в аспекте биотеррористической настороженности последних лет и эта версия кажется весьма привлекательной. Обе версии, несмотря на внешнюю эффектность, все же игнорируют ряд весьма важных экспериментальных наблюдений последних лет, что в конечном итоге уводит исследователей от понимания черной смерти как сложного природного процесса, способного повторяться в виде пандемической катастрофы.
Чума - как природно-очаговый сапроноз
Учение о природной очаговости чумы, разработанное в начале ХХ столетия, было построено на положении о первичности животных (различные виды грызунов), как резервуара возбудителя чумы. При массовой активизации таких очагов в разгар третьей пандемии чумы , это учение позволяло вполне убедительно связывать возникновение вспышек чумы в населенных пунктах с распространением больных чумой грызунов. Однако по мере угасания природных очагов чумы, еще пока еще не объясненном процессе, начавшемся в конце 1930-х гг., учение все чаще переставало работать . Чума не появлялась там, где, казалось, она должна обязательно быть, но возвращалась там, где ее не ждали. Последнее внешне проявлялось одномоментным развитием на больших территориях чумных эпизоотий среди диких грызунов, однако, тщательный контроль над таким очагом, проводившийся десятилетиями до этого, свидетельствовал о том, что чумы все эти годы в нем не было. Причины инфицирования грызунов возбудителем чумы оставались неясными, да и роль самих грызунов в поддержании возбудителя чумы в природе становилась все менее ясной по мере расширения масштабов таких исследований.
Под давлением эпидемиологических наблюдений, Mollaret H. (1963), первым среди чумологов, был вынужден вернуться к забытому в начале XX столетия учению Макса Петтенкофера (1885), предполагающему участие почвы в поддержании в природе возбудителей опасных инфекционных болезней. Смысл его гипотезы сводится к тому, что чумной микроб при наличии соответствующих условий может длительно персистировать в почве нор грызунов ( теллурическая чума ). Развивая гипотезу Mollaret, М. Балтазар (1964) пришел к заключению, что цикл чумы в природных очагах состоит из двух фаз: паразитической (на грызунах и их блохах - кратковременный и неустойчивый) и непаразитарной (существование в почве нор - устойчивый). Механизм данного явления оставался не понятным до тех пор, пока С.В Никульшин. с соавт. (1993) не показали, что ряд амеб способны фагоцитировать Y.pestis и сохранять ее в предцистах.
А.И. Дятлов (1989), Сунцов и Сунцова (2000), последовательно придерживающиеся антропоцентрических воззрений на патогенность Y.pestis, привели весьма интересные наблюдения в пользу ее исключительно малого знакомства со своими теплокровными жертвами: возбудитель чумы не способен к гемолизу эритроцитов крови, т.е. он не является паразитом крови; у него отсутствуют приспособления к адгезии к клеточным мембранам хозяина (если им считать человека), а, следовательно, в процессе коэволюции теплокровных и возбудителя чумы (если ее тоже допустить) не совершенствовалась система тканевой комплементарности; отсутствует органотропность; возможность трансмиссии к другим хозяевам посредством эктопаразитов появляется только после развития сепсиса - неспециализированной формы инфекционного процесса и одновременно его терминальной стадии; температурная зависимость антигенных свойств; способность заражать разные виды животных (отсутствие видовой специализации); нестабильная способность к образованию блока преджелудка блох и отсутствие у блох физиологических механизмов предотвращения блокообразования или освобождения от блока; блокированные блохи быстро гибнут в результате обезвоживания и голодания; блохи инфицированные, но освободившиеся от блока имеют значительно более короткий срок жизни, чем неинфицированные (такова эволюционная цена, заплаченная Y.pestis за оперон hms). Уже по этим данным можно предположить, что во время чумных вспышек и пандемий, сколь масштабными они не были бы, человек не более чем случайный хозяин чужого паразита.
Противоречат представлениям о существовании в природе чумного микроба как облигатного паразита теплокровных животных, и данные сравнения геномов Yersinia pseudotuberculosis и Y.pestis. Эти исследования осуществляются на основе предположения о том, что псевдотуберкулезный микроб является предком возбудителя чумы (В.В. Сунцов и Н.И. Сунцова, 2000; U. Vogel и H. Claus, 2000). Однако интерпретация их результатов для ученого, привыкшего считать, что патогенность микроорганизмов обусловлена приобретением генов факторов патогенности, требует преодоления некоторого психологического барьера. Возбудитель чумы, более патогенный для человека и большинства модельных животных, чем возбудитель псевдотуберкулеза, утрачивает значительную часть генов, которые традиционно относят к генам вирулентности и патогенности. По данным, приведенным А.П. Анисимовым (2002), Y.pestis утрачивает гены адгезинов, уреазы (сдвиг рамки считывания), инвазинов Inv и Ail (вставка IS-элементов), подвижности, способности к синтезу О-боковых цепей ЛПС (не установленные механизмы образования мутаций) и ряд других. Из 17 биосинтетических генов, выявленных у псевдотуберкулезного микроба, 5 в геноме Y.pestis инактивированны за счет вставок и делеций. Компьютерный анализ полного генома чумного микроба (штамм CO92) показал наличие 149 псевдогенов. Эволюционные же приобретения чумного микроба весьма сомнительны, если его рассматривать как облигатного паразита человека и теплокровных животных. Это плазмида рPst - кодирует пестицин Р1, являющийся бактериоцином, блокирующим рост псевдотуберкулезного микроба серотипа 1 и штаммов Y.pestis, не образующих Р1; а так же фибринолизин и плазмокоагулазу - отсутствуют у полевочьих штаммов Y.pestis, т.е. для поддержания чумного микроба среди грызунов эти три фактора не обязательны; и плазмида pFra - кодирует капсульный антиген и мышиный токсин , их значение, как факторов вирулентности чумного микроба для теплокровных организмов, в последние годы подвергается сомнению (см. у Домарадского, 1998). В число таких приобретений входит и конститутивная экспрессия гена hmsT, составной части оперона hms, обеспечивающего образование блока в преджелудке блохи. Здесь уместно отметить, что аналогичное явление - упрощение генома в обмен на увеличение специализации и вирулентности, наблюдается у других опасных для человека микроорганизмов, например, у возбудителей туберкулеза и проказы (Cole S.T., Buchrieser С., 2001), вирусов натуральной и обезьяньей оспы (Маренникова С.С., Щелкунов С.Н., 1998). Ранее нами (Супотницкий М.В., 2000) было высказано предположение, что, данный феномен является проявлением того же механизма дегенеративной эволюции, который использует естественный отбор для развития видовой специализации у многоклеточных паразитических организмов (например, специализация к человеку у ленточных червей приводит к утрате ими пищеварительной системы и к упрощению до предела нервной).
Приведенные выше данные позволяют сделать предположение о том, что Y.pestis, являясь дегенеративным производным имеющего значительно более широкий круг хозяев псевдотуберкулезного микроба, в организме человека и грызунов проявляет вирулентность в виде локального феномена - способности пролиферировать в клетках белой крови. При этом Y.pestis использует те же механизмы специализации, которые позволяют ей поддерживаться среди почвенных одноклеточных организмов, эволюционных предшественников фагоцитов. Так как жизнь или смерть случайно инфицированного теплокровного организма ничего не значит для поддержания такого паразита в природе, то его вирулентность не лимитируется необходимостью сохранения жизни своим жертвам. Болезнь протекает в септической форме, а массовые инфекции носят характер вспышек, и редко напоминают классический эпидемический процесс, т.е., передачу возбудителя чумы от одного заболевшего к другому (только в отдельных случаях легочных осложнений также носящих случайный характер; см. в следующем сообщении).
То обстоятельство, что устойчивое сохранение возбудителя чумы в природе обеспечивается, главным образом, почвой, где Y.pestis существует в некультивируемом виде как облигатный паразит одноклеточных организмов, принципиально меняет методологию изучения аспектов эпидемиологии чумы, имеющих отношение как к механизмам поддержания в природе возбудителя болезни, так и к его проникновению в человеческие популяции. Инфицированные грызуны должны рассматриваться не как природный резервуар чумы, а как индикатор сдвига в экологии одноклеточных организмов - естественных хозяев Y.pestis. Те территории, на которых возбудитель чумы поддерживается эпизоотиями среди грызунов, следует рассматривать, как вершину айсберга чумы , ставшую хорошо различимой благодаря применению методов медицинской бактериологии. Границы же его подводной , наиболее опасной части, еще только предстоит установить методами молекулярной биологии.
Нарушение экосистемы, вмещающей чумной микроб, приводит к выходу Y.pestis за ее пределы (почва нор, сосудистые растения) и к инфицированию грызунов и их эктопаразитов. Такое явление возможно в результате сложно опосредованных климатических воздействий и глобальных катастрофических процессов, изменяющих абиотические (микроэлементный состав почвы, ее температура, влажность, рН, содержание О2 и т.п.) и биотические факторы (наличие пищи, пищевых конкурентов и хищников, освоение новых экотопов и др.), поддерживающие экосистемы простейшие - Y.pestis . Поэтому оно наблюдается крайне редко, скорее как исключение, чем закономерность.
Сама же эпидемиология чумы нуждается в новых определениях, отражающих принципиальные изменения в понимании механизмов ее существования в природе, и позволяющих описывать чуму как эпидемическое явление, но вне учения о природной очаговости чумы начала ХХ столетия.
Природный резервуар возбудителя чумы - совокупность одноклеточных организмов - биологических хозяев Y.pestis, без которых ее существование в природе как биологического вида невозможно (фактор Y в понимании Макса Петтенкофера).
Природный очаг чумы - географический ландшафт, в почве которого методами молекулярной диагностики доказано присутствие возбудителя чумы в некультивируемом состоянии (холодный очаг), и/или на его территории фиксируются эпизоотии и эпидемии чумы (пульсирующий или активизировавшийся очаг).
Реликтовый очаг чумы - территория, неопределенно долго включающая природный очаг чумы, о существовании на которой в прошлом вспышек чумы среди людей известно из исторических источников.
Усилители природного резервуара чумы - биотические объекты, не имеющие значения для поддержания в природе возбудителя чумы как биологического вида, но способные накапливать, размножать и доставлять его в организм определенного вида теплокровных животных или человека.
Активизировавшийся природный очаг чумы - продолжающееся в течение исторически зафиксированного периода времени (до нескольких столетий) появление на территории природного очага чумы, чумных эпизоотий и эпидемий.
Пульсация природного очага чумы - кратковременное (до нескольких лет), интенсивное, охватывающее обширные территории, проникновение возбудителя чумы из его природного резервуара в популяции диких и домашних животных и их эктопаразитов.
Цикл глобальной активизации чумы - скачкообразное нарастание колебательных процессов в экосистемах, включающих чумной микроб, и проявляющееся пульсациями природных очагов чумы (отдельные эпизоотии и вспышки чумы среди людей в регионах, ранее считавшиеся от нее свободными), сменяющееся после прохождения пика (пандемия чумы) их постепенным угасанием (период упадка чумы - современное состояние с заболеваемостью чумой в мире).
Этими определениями мы будем пользоваться при дальнейшем изложении материала.
"Черная смерть"
Второй пандемии чумы предшествовали глобальные климатические и тектонические катаклизмы. С началом малого ледникового периода (XIII столетие), климат стал холоднее, неустойчивее, сократился вегетационный период растений. Период 1270-1350 гг. характеризуется увеличением внутрисезонной изменчивости климата. Однако появление чумы среди людей нельзя связать с расширением степных зон, и, соответственно, увеличением полевых грызунов. Погода на юге Европы в течение нескольких лет перед черной смертью отличалась теплом и сыростью. В 1342 г. - обилие снега зимой и сильные дожди летом - поля Франции опустошены сильным наводнением, в Германии затоплено много городов. В 1343 г. отмечены постоянные дожди и наводнения. С 1345 г. по всей Европе период особенной сырости , продолжавшийся еще несколько лет, постоянные неурожаи, нашествия саранчи до Гольштинии. К концу столетия льды полностью загородили путь в Гренландию (Гезер, 1867; Борисенков, Пасецкий, 1988).
Распространение пандемии черной смерти. Попробуем хотя бы приблизительно установить границы реликтовых очагов чумы, с пульсации которых началась пандемия черной смерти. Для европейцев чума началась в 1346 г. на побережье Черного моря и в устье Дуная. Византийский историк Никифор (Nicephorus) так изложил эти события: Около этого времени овладела людьми тяжкая чумоподобная болезнь, которая, двигаясь от Скифии и Меотии, и от устьев Дуная, господствовала еще в первую весну. Она оставалась весь этот год (1347), проходя только в точности по берегам, и опустошала как города, так и села, и наши, и все, которые последовательно простираются до Гада и Столбов Геркулесовых. В следующем году она отправилась и к островам Эгейского моря; потом поразила Родос, так же Кипр и жителей остальных островов (цит. по Гезеру, 1867).
В записи, датированной 1346 г., русский летописец свидетельствовал: Того же лета казнь была от Бога на люди под восточною страной на город Орнач (при устье Дона - Карамзин) и на Хавторокань, и на Сарай и на Бездеж (город на рукаве Волге, ниже Енотаевки) и на прочие грады во странах их; бысть мор силен на Бессермены (хивинцы) и на Татары и на Ормены (армяне) и на Обезы (абазинцы) и на Жиды и на Фрязы (генуэзцы и венецианцы в колониях при Черном и Азовском морях) и на Черкасы и на всех тамо живущих (Воскр. лет., стр. 210; цит. по Рихтеру А., 1814).

Деталь иллюстрации «Похороны жертв чумы в Турне» (« Burying Plague Victims of Tournai »). Взята из рукописи, называемой «Хроники Гилля Майзета» (« Annals Gilles de Muisit », 1272 – 1352 гг.), настоятеля монастыря святого праведника Мартина. Показанные художником события относятся к чуме «черной смерти» в бельгийском городе Турне в 1349 г. Рукопись хранится в Библиотеке короля Альберта Первого, Брюссель.
Следовательно, в 1346-1347 гг. на территории, включающей низовья Волги, Северный Прикаспий, Северный Кавказ, Закавказье, Крым, Восточные отроги Карпат, Причерноморье, Ближний и Средний Восток, Малую Азию, Балканы, Сицилию, Родос, Кипр, Мальту, Сардинию, Корсику, Северную Африку, юг Пиренейского полуострова, устье Роны, синхронно началась пульсация природных очагов чумы. Огромные количества Y.pestis посредством инфицированных эктопаразитов вбрасывались из ее природного резервуара в человеческие популяции. Произошла столь массовая гибель людей, что умерший от чумы человек вызывал столько же участия, сколько издохшая коза (Боккаччо Дж., 1351).
Клиника черной смерти
По меткому замечанию Гезера Г. (1867), черная смерть если и уклонялась от обычной картины чумы, то это лишь потому, что она совмещала в себе все те явления, которые были порознь в различных чумных эпидемиях. Прежде всего, авторами, современниками черной смерти, выделялся синдром febris continuae - непрерывная лихорадка . Под ним понимался следующий комплекс симптомов: черный и сухой язык, бред и взрывы бешенства, чувство тоски и боли в стороне сердца, ускоренное дыхание, кашель, разного рода мокроты, мутная и нередко черная моча, черные испражнения на низ, черная кровь. На фоне febris continuae появлялись петехии, карбункулы и бубоны. Трупы людей, умерших от чумы, к всеобщему ужасу живых, стремительно чернели и напоминали по цвету уголь - отсюда и появилось название черная смерть .
Такой клинический симптом при чуме, как кровохаркание, не был известен врачам из прежней практики. Шольяк называл черную смерть чумой с кровохарканьем. Как правило, такая форма болезни появлялась в начале эпидемического процесса, локально, безо всякой связи с эпидемиями чумы в других городах, затем она сменялась бубонной формой. Шольяк так же отмечал значительно большую заразность легочной формы чумы по сравнению с бубонной.
Однако не все ученые того времени придерживались его точки зрения на заразность легочной чумы. Последовательный контагионист Ибнулкатиб утверждал, что многие люди, не смотря на контакты с больными с такой формой болезни, оставались здоровыми, и наоборот, получали ее вообще безо всяких контактов, и не будем спешить отвергать его наблюдения. Не прошло и ста лет, упоминания в летописях о чуме с кровохарканием , прекратились
Характер эпидемий
В восприятии современников (Ковино; цит. по Гезеру, 1867), появлению черной смерти предшествовало какое-то отравление, вызванное теллурическими факторами, например парами, поднимавшимися из расщелин земли. Внешне это проявлялось следующим образом. Перед эпидемией в каком-нибудь городе обычно наблюдали людей, у которых отмечали бледность лица и болезненные ощущения в паховой области; этот факт и интерпретировали как отравление парами . Если следовать современным представлениям о распространении чумы, то появление малоболезненных бубонов означает то, что задолго до появления чумы в клинически выраженной форме она проникала в отдельные группы населения с маловирулентными штаммами Y.pestis. В начале эпидемии появлялось большое количество людей с так называемой амбулаторной чумой (pestis ambulans), но так как их за больных тогда не считали, то и начало самой эпидемии не замечали. Поэтому черная смерть в населенном пункте обычно начиналась взрывом смертельных случаев болезни, точнее, обращала на себя внимание после такого взрыва. Так, в Авиньоне, в январе 1348 г., чуму обнаружили только после того, как все монахи одного монастыря (около 700 человек) умерли в одну ночь. Затем эпидемия с необычной быстротой распространялась среди жителей населенного пункта и, поглотив огромное число жертв, она заканчивалась обычно через 5-6 месяцев.
Во время эпидемии черной смерти сначала заболевали дети, затем женщины, затем уже остальное бедное население. Когда чума возвращалась (как правило, в бубонной форме), то она истребляла людей богатых, совсем не пострадавших во времена ее первого появления (Ковино).
Черная смерть продолжалась до начала 1352 г. и унесла, по некоторым оценкам, до трети населения Земли. Остаток XIV века и в начале XV века, чума ежегодно появлялась в Европе и в России то в одном, то в другом месте. Однако ее удары постепенно ослабевали.
Хронология распространения черной смерти в Европе в 1346-1350 гг.*
|
Год |
Месяц |
Страна или местность |
|
1346 |
— |
Сицилия, некоторые местности в Италии (?), Константинополь |
|
1347 |
Весна, первая половина года |
Константинополь, Кипр, Греция, Мальта, Сардиния, Корсика, некоторые прибрежные города Италии |
|
Начало ноября |
Марсель |
|
|
1348 |
Начало года |
Испания (Альмерия), Южная Франция (Авиньон) |
|
Март |
Нарбонна, Модена |
|
|
Весна |
Флоренция — Лангедок, Гасконь, Франш-Коле |
|
|
Май |
Валенция (Испания) |
|
|
Июнь |
Паченца, Падуя, Валенция (наибольшее развитие эпидемии), Барселона — Перуджия |
|
|
Август |
Рим, Южная Англия, Шлейзвиг — Гольштейн |
|
|
Сентябрь |
Сарагоса (наибольшее развитие эпидемии) |
|
|
Ноябрь |
Лондон — Норвегия (Берген) |
|
|
Декабрь |
Далмация — Ютландия |
|
|
1349 |
Январь |
Каринтия |
|
Весна |
Вена, Франкфурт на Майне |
|
|
Конец мая |
Прекращение эпидемии в Англии |
|
|
Август |
Любек, Шлезвиг, Торн |
|
|
1350 |
Лето |
Польша |
*По Г. Гезеру (1867), ссылавшегося на недоступную для нас работу Геккера (Hecker J.F.C.,1838).
Откуда взялась версия о применении татарским ханом БО
В основе этой версии, далеко не безобидной для подозреваемых в разработке биологического оружия стран Третьего мира, лежит всего на всего единственное свидетельство нотариуса Габриэля де-Мюссе, пережившего чуму в городе генуэзских работорговцев - Кафе (современная Феодосия). Впервые оно была опубликовано в 1842 г. Геншелем по ранее не известной рукописи, хранившейся в Бреславле, в Гедигеровской библиотеке. Судя по клинической картине, подробно описанной де-Мюсси, среди осаждавших город татар и его защитников, преобладала бубонная форма чумы, распространившаяся из вторичных крысиных очагов посредством инфицированных блох. На ее фоне развивались вторичные легочные осложнения, но они не были доминирующими для этой эпидемии. Однако историки медицины обращают свое внимание, прежде всего, на следующее замечание де Мюсси: Родные, друзья и соседи поспешили к нам, но мы принесли с собой убийственные стрелы, при каждом слове распространяли мы своим дыханием смертельный яд . Отсюда делается вывод о распространении легочной чумы здоровыми носителями, прибывшими из Кафы, и упорно замалчиваются свидетельства о бубонных формах болезни (хотя бы того же Боккаччо, 1351), имевших место во всех европейских городах. Причина эпидемиологической путаницы тут в том, что современные авторы, средневековые и, даже, XIX столетия, описывая одни и те же события, используют разную научную терминологию.
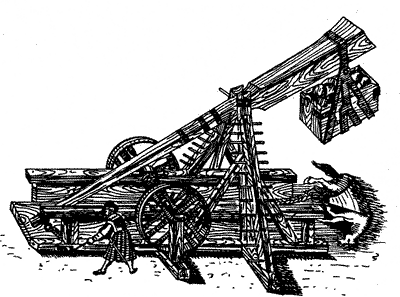
Забрасывание в осажденный город мертвой лошади с помощью метательной машины. По сути — это биологический террор на основе миазматических представлений о распространении эпидемических болезней. С рисунка Леонардо да Винчи (1445—1520).
Для де-Мюсси (1350) убийственная атмосфера это загнившая пневма , так как пользовался он пневматическими представлениями о распространении даже не контагия чумы, а еще миазмов в понимании Авиценны, Галена и Гиппократа. Иного объяснения этих событий ни он, ни кто другой в эти годы просто не воспринял бы, - до книги Фракасторо О контагии, контагиозных болезнях и лечении еще должно было пройти 200 лет. Однако на момент опубликования его записок, учение Фракасторо было доминирующим в понимании распространения чумы. В соответствии с ним, единственная причина чумы - специфический яд (контагий), который выходит вроде испарины из тела больных, распространяется в воздухе только на самых малых расстояниях; но взамен того, прилипает ко всем вещам и предметам; что некоторые из них могут сохранять в себе чумной яд до тридцати и больше лет и, следовательно, такие вещи и предметы могут быть переносимы на безграничные пространства, и послужить к заражению целых городов . Современные авторы понимают под заносом здоровыми людьми их способность распространять вокруг себя бациллы чумы.
Бубонная же чума не выходит за пределы своего природного очага, следовательно, генуэзцы не могли ни заразить всю Европу (процесс распространения чумы продолжался 5 лет), ни поддерживать эпидемии чумы до первой половины XIX столетия. Де-Мюссии, чудом вырвавшийся осенью 1348 г. из одного природного очага (Северного Причерноморья - см. ниже) и попав в другой (Реликтовые очаги чумы долины реки По), не знал, что чума уже распространяется по Южной Европе и её Средиземноморскому побережью (см. таблицу.). Поэтому у него сложилось такое представление, что где бы не приставали итальянские корабли, везде быстро умирали все те, кто соприкоснулся с прибывшими на них. Почти генетический страх перед чумой, выработанный столетиями ее беспощадного появления, мешает перейти нам рубеж понимания того, что она ниоткуда не заносится и никуда не исчезает .
Возвращение легочной чумы
Первое свидетельство о возвращении легочной чумы относится к 1815 г., когда в индийской провинции Куче и на юго-востоке провинции Гузерат (штат Гуджарат) появилась смертельная болезнь с кровохарканием и бубонами. Болезнь продолжилась на следующий год, и с января по март 1816 г. сильно проредила местное население (Гирш, 1857). В России легочная чума, впервые с XV столетия, напомнила о себе в станице Ветлянская в 1878 г.
По принятой сегодня классификации (Руднев Г.П., 1940), обе вспышки были вторично-легочной чумой. Но вот любопытное отличие, не позволяющее их полностью отождествлять с черной смертью. Никто из врачей не наблюдал карбункулов, и лишь очень немногие видели петехии, тела погибших не казались обугленными.
Крайне интересны результаты поиска аналогичных эпидемий, проведенные Гиршем (1857) в трудах авторов того времени. Оказалось, что в описаниях эпидемий чумы, распространявшихся с ХVI по XIX столетие, нет и речи о кровавом кашле, как о клиническом проявлении чумы. Только возвратясь к XIV столетию Гирш обнаружил в трудах европейских ученых и русских летописцев описания легочной чумы в пандемическом распространении .
Реликтовые очаги чумы
Доказательства существования Y. pestis в одноклеточных организмах в принципе меняют и так довольно запутанные представления о географическом распространении чумы. Попытки определить ее древние природные очаги в Европе делались неоднократно, однако, в их основе лежали представления о первичности животных (различные виды грызунов), как резервуара возбудителя чумы. Чтобы избежать упреков со стороны оппонентов в отсутствии научного подхода, исследователи, очерчивая границы таких очагов, старались состыковывать исторические сведения с ареалами тех или иных грызунов, появление же чумы за их пределами объяснялось заносом . Например, Миронов Н.П. (1958), проанализировав ископаемые останки флоры и фауны, сделал вывод о том, что северная граница древнего очага чумы проходила через Балканы по линии Киев-Нижний Новгород . Федоров В.Н. (1960) определял ее на основании анализа сведений о локальных вспышках чумы в Европе в позднее, чем черная смерть, время (XVI-XVII вв.), так как тогда границы очагов вписывались в ареал крапчатых сусликов. Но исторические источники свидетельствуют о значительно больших территориях Европы, периодически становящихся эндемичными по чуме. Для приблизительного определения их границ мы воспользовались работами Рихтера А. (1814), Гезера Г. (1867), Архангельского Г.Ф. (1879), Щепотьева Н.К. (1897), Ахшарумова Д.Д. (1900), Дербека Ф.И. (1905), Васильева К.Г. и Сегала А.Е. (1960), Козлова М.П. и Султанова Г.В. (1993). Для систематизации реликтовых очагов мы использовали синхронистический принцип. Точные же границы таких очагов можно определить только с помощью методов молекулярной диагностики.
Реликтовый Северо-западный природный очаг чумы. Представляет собой восточную оконечность цепочки очагов чумы, тянущихся от побережья северной Франции (пролив Ла-Манш), через территории Голландии, Дании, германских государств, Польшу, Прибалтику и по Карелии до южного побережья Белого моря. Активизация очага зафиксирована в русских летописях XIII столетия и приходится на начало малого ледникового периода. В 1424 г., судя по территории, охваченной чумой, пульсация очага достигла максимума - бубонная и легочная чума в Новгороде, Пскове, Карелии, а также в Литве и Немцах . Последняя эпидемия чумы в Пскове пришлась на 1711 г., в Новгороде на 1606 г.
Реликтовый природный очаг чумы Русской равнины. Включает территорию, по форме приближающуюся к четырехугольнику с углами Великие Луки-Вологда-Кострома-Смоленск. В 1229-1230 гг. активизация очага чумы дала о себе знать мором в Смоленске, погибло до 32 тыс. человек.
В 1654-1656 гг. пульсация очага достигла максимум. В Москве эпидемия началась в августе 1654 г. и продолжалась до января 1655 г., погибло до 300 тыс. человек. С августа по сентябрь 1654 г. чумой были охвачены Кострома, Тверь, Коломна, Торжок, Звенигород, Кашин, Переславль-Рязанский, Переславль-Залесский, Суздаль, Тула, Калуга, Углич, Нижний Новгород, Казань, многие их слободы и уезды. Осенью 1771 г. в Москве и ближайших к ней городах разгорается эпидемия чумы, сопровождающаяся бунтом населения. Однако по охвату территории ее масштабы были значительно меньшими, чем в 1654 г. В 1773 г. пульсация очага прекратилась.
Реликтовые природные очаги чумы Северного Причерноморья и Малоросии. Это восточная оконечность последовательности реликтовых очагов чумы, тянущихся от Балкан на восток вдоль всего северного побережья Черного и Азовского морей до очагов чумы Великого Евразийского чумного излома (см. ниже). На севере границы очага можно расположить на равнинном пространстве между Волыно-Подольской и Средне-Русской возвышенностями до границ Курской области. В 1738 г. активизация очагов чумы Северного Причерноморья и Малоросии достигла максимума. Во второй половине XVIII столетия чума постепенно отступает в направлении с севера на юг и дробится на мелкие очажки. В первой половине XIX столетия активными остаются только очаги чумы Северного Причерноморья, расположенные на пространстве между Крымским полуостровом и Тирасполем, причем их границы постепенно сужаются.
Великий Евразийский чумной излом. Под ним мы понимаем последовательность реликтовых и пульсирующих природных очагов чумы, расположенных изломанной дугой - от пустынь Йемена до Иранского нагорья (Саудовско-Йеменский очаг с центром в Эль-Асире и Хавлане, Сирийско-Месопотамский пустынный очаг, Курдо-Иранский природный очаг чумы, очаги чумы Центрально-Иранского плоскогорья, Северо-Иранских краевых гор, а также смежных с ними плоскогорий и впадин Афганистана), затем через Кавказский хребет (Приараксинский участок очаговости чумы, Закавказский равнинно-предгорный очаг чумы, Закавказский высокогорный природный очаг чумы, Центрально-Кавказский природный очаг чумы, Восточно-Кавказский природный очаг чумы), Прикаспийскую низменность (Прикаспийский северо-западный очаг чумы), Зауралье (Зауральский природный очаг чумы), равнины среднего Поволжья (Волго-Уральский природный очаг чумы), долины рек Ветлуга, Вятка и Кама до Северных Увалов (реликтовые очаги). В пульсации этих очагов прослеживается определенная пространственная и временная последовательность. Сначала чума опустошает Месопотамию, Персию и Малоазиатскую часть Турции, затем, в течение двух-трех десятилетий, она продвигается с юга на север, и, описав дугу вокруг Каспия со стороны Кавказского хребта, ударяет по Астрахани (1662, 1693, 1727, 1806, 1877), после чего поднимается по Волге, иногда до Саратова (1808), реже ещё дальше (Нижний Новгород, 1363; Казань, 1665, Вятка, 1667).
Современники черной смерти зафиксировали активизацию очагов чумы Великого Евразийского чумного излома - от Курдо-Иранского, до Прикаспийского Северо-западного. В их восприятии это была другая волна чумной эпидемии (Гезер Г., 1867), прокатившаяся через Сирию, Армению, Малую Азию и прикаспийские земли.
На территории России на которой в 1346 г. наблюдались пульсации реликтовых очагов чумы, наиболее северным участком Излома, видимо, была местность в районе Енотаевки (современная Астраханская область), где по указанию летописца произошла казнь от Бога на жителей города Бездеж (Воскр. лет.). Самой южной территорией Излома с пульсирующими очагами времен черной смерти, можно считать местность между реками Тигр и Евфрат в районе Багдада (1347), охваченную бубонной и бубонно-септической чумой с ужаснувшей современников повальной смертностью населения.

Чума на территории реликтового северо-западного очага чумы. Одно из клейм иконы «Видение пономаря Тарасия», с изображением «Симеоновского мора» в Новгороде в 1467 г. (из книги Н.А Богоявленского,1960)
В годы последующих пульсаций Излома , территории пульсирующих очагов расширялись и смещались на север и на юг. В 1363 г. границы пульсирующих очагов поднялись до Нижнего Новгорода, ...а пришел (мор) от низу, от Бездежа, ...и опусте земля вся и порасте лесом, и бысть пустыни всюду непроходимы (Никоновская летопись, с. 3) . До средины XVII столетия о чуме на территории Излома в исторических источниках не упоминается, возможно, это не от невнимательности летописцев. Например, Петр Петерей писал: Московиты, находящиеся за Рязанью и в Татарии, вовсе не знают моровой язвы, страдают же от нее лишь близкие к западной границе, а именно: Новгород, Псков, Смоленск и др. (История о Великом княжестве Московском, 1620).
Пульсации очагов Излома возобновились в средине XVI столетия, и их максимум на территории Европы совпал с наибольшей активностью Реликтового природного очага чумы Русской равнины и большинства других европейских реликтовых очагов. Летом 1655 г. моровое поветрие появилось в низовьях Волги и в Астрахани. Затем эпидемия вспыхнула Казани, где по данным С.М. Шпилевского (1878), в тот год от чумы погибло 48 тыс. человек. В 1657 г. чума свирепствовала и в соседней с Казанской - Вятской области (северная граница Излома ), и в самом городе Вятке. Летом 1657 г. чума снова возобновилась в низовьях Волги, в 1692 г. чума повторилась в Астрахани.
Южная граница реликтовых очагов Излома проходит по побережьям Персидского залива (на восточном побережье, по крайней мере, достигает Бушира). Максимум в их активности проявился чудовищными эпидемиями чумы в Багдаде и Басре в 70-х гг. XVIII столетия (до 2-х млн. погибших). В последующие годы северные территории пульсирующих очагов Излома (Россия), сужались и смещались к югу (сегодня это Прикаспийские, Волго-Уральские и Кавказские очаги чумы), южные (Месопотамия), наоборот, подтягивались на север (Закавказские и Иранские очаги чумы). Количеств жертв чумы в одних и тех же природных очагах снижалось от вспышки к вспышке, к средине 50-х гг. XIX у ученых появилось иллюзорное представление об окончательной победе над чумой.
Балкано-Придунайские реликтовые очаги чумы. Синхронно активизировались как в первую, так и во вторую пандемии чумы, приводя к массовой гибели людей. Среди них можно выделить три группы реликтовых очагов: 1) цепочка очагов на равнинной местности между реками Прутом и Серетом, расположенных от Хотина до Измаила (включая Яссы, Браилов, Галац и др.), своей восточной частью вплотную прилегающих к реликтовым очагам Северного Причерноморья и Малороссии; 2) цепочка очагов на равнинной местности между Дунаем и Черноморским побережьем от Бабадага до Варны (включая Черноводы, Каварну, Кюстенджи); 3) отдельные очаги на равнинной местности между Константинополем, Андрианополем и побережьем Мраморного моря (проявляли небольшую активность так же и в третью пандемию чумы). Максимум активности достигнут в конце XVIII столетия.
Реликтовые очаги чумы долины реки По. Исторически они наиболее смертоносные в Европе и самые активные на Апеннинском полуострове; многократно разгорались в первую и вторую пандемии. Их пульсациями объясняются сокрушительные эпидемии Средневековья и эпохи Возрождения в городах Венеция, Милан, Падуя, Болонья, Пьяченца, Палермо, Верона и др., иногда воспринимаемых современниками как конец света . Максимум активности достигнут в средине XVII столетия.
Реликтовые очаги чумы долины реки Арио. Катастрофические эпидемии чумы в Пизе и Флоренции во времена первой и второй пандемий, пик активности приходится на времена черной смерти (художественное описание см. у Боккаччо Декамерон ).

«Чума в Перуджии в 1464 г.» Фрагмент фрески итальянского художника Бенедитто Бонфигли (Benedetto Bonfigli ; 1420-1496). Ангел атакует демона смерти, поражающего горожан стрелами чумы. Перуджия — главный город одноименной итальянской провинции.
Реликтовые очаги чумы долины реки Вольтурно. Их пульсации проявлялись тяжелыми эпидемиями в Неаполе в период второй пандемии чумы. Самая смертоносная эпидемия чумы вспыхнула в 1656 г. Она унесла жизни 200 тыс., а возможно и 270 тыс. человек из 400 тыс. населения города Последний раз активизировались в 1900 г.
Реликтовые очаги чумы долины реки Рона. По многочисленным историческим источникам очень активны. Пульсации проявлялись сокрушительными эпидемиями чумы во времена первой и второй пандемий. Границы очагов можно заключить в треугольник Авиньон (вершина) - Тулон и Монпелье (основание) . Максимум активности достигнут в 1720-1722 гг.
Сицилийские реликтовые очаги чумы. Располагаются вдоль северного побережья острова между городами Трапани и Сиракузы. Наиболее активно пульсация очагов происходила в период второй пандемии на местности, прилегающей к городу Мессина.
Реликтовые очаги чумы Пиринейского полуострова. Долина реки Дуэро (местность в районе города Порто); долина реки Гвадалквивир (Севилья); местность южнее и восточней Андалузких гор, включающая Гибралтар, Малагу, Альмерию; низовья реки Турия (Валенсия); долина реки Эбро (Сарагоса); местность, прилегающая к Каталонским горам с востока (Барселона). Пик активности приходится на средину XVII столетия.
Реликтовые очаги чумы долины реки Гаронна. Пульсации проявлялись сокрушительными эпидемиями чумы в городах Бордо и Тулуза во времена второй пандемии. Пик активности приходится на конец XVII столетия.
Реликтовые очаги чумы северо-восточной Франции. Это обширное равнинное пространство между городами Руан, Амьен, Реймс и Париж, включающее долины рек Сена, Марна, Сомма. Мощные пульсации во времена первой и второй пандемий (пик активности приходится на средину XVII столетия).
Реликтовые очаги чумы полуострова Корнуолс (о. Великобритания). Равнинная местность по форме напоминающая неправильный параллелограмм, расположенная между Бристолем и Уэтмутом, и вдоль юго-восточного склона Корнийских гор, где в августе 1348 г. вымерло почти все население ( люди ложились, подобно колосьям под серпом жнеца ).
Реликтовые очаги чумы Юго-Восточной Англии. Расположены на местности, включающей Лондон и его пригороды в радиусе 20 миль от Сити. Пульсации достигли максимума в средине XVII столетия.
Реликтовые центрально-европейские очаги чумы. Цепочка реликтовых очагов, расположенных вдоль северных отрогов Альп (сокрушительные эпидемии чумы второй пандемии в городах Женева, Базель, Берн, Цюрих, Мюнхен, Линц, Вена), и соединяющихся с западной оконечностью Балканских реликтовых очагов чумы (пик активности приходится на первую половину XVII столетия).
Реликтовые очаги чумы долин рек Рейн, Везер, Эльба. Во время второй пандемии их эпидемическая активность нарастала по мере приближения к побережью Балтийского моря (сокрушительные эпидемии чумы второй пандемии в Страсбурге, Франкфурте, Кельне, Ганновере, Гамбурге, Магдебурге и др.). Видимо сливаются с цепочкой реликтовых очагов чумы, расположенных вдоль побережья пролива Ла-Манш, Северного и Балтийского морей. Пик активности приходится на первую половину XVII столетия. Наиболее упорно эпидемии чумы держались на местности, включающей в себя города Дрезден (1680), Магдебург (1681), Галле (1682), Гальберштад, Брауншвейг, Эрфрут, Нордгаузен и Мюльгаузен (1682-1683) - Германское чумное пятно . На его территории отдельные вспышки и случаи чумы встречались до 1711 г., после чего она совершенно исчезла из Германии.
В конце XV столетия, среди многократно отсортированного чумой и натуральной оспой населения европейских городов, вспыхивают тяжелые и упорные эпидемии сифилиса. Для людей оставшихся генотипов было характерным необычайно раннее наступление второй стадии болезни (вторичный сифилис на 9-20-й день появления клиники болезни); резкие боли в суставах, доводящие больных до отчаяния; язвы с глубоким распадом мягких тканей; поражение костей через год болезни и образование гумм уже через полтора года. Такой сифилис был воспринят населением как новая болезнь .
Отступление чумы из Европы
Оно происходило в направлении с севера на юг, т.е. в обратном ее распространению в 1346-1351 гг. В Соединенном Королевстве последняя эпидемия чумы пришлась на 1665 г. ( Великая Лондонская чума ), в Германии на 1711 г. Последняя крупная эпидемия чумы в юго-западной части континента вспыхнула в 1721 г. в Провансе, на территории реликтовых очагов долины реки Рона. В начале 1830-х гг. в Восточной Европе погасли Балкано-Придунайские реликтовые очаги чумы. С 1840-х гг. перестали напоминать о себе гибелью людей Кавказские очаги чумы. Прекращение чумных эпидемий ученые объяснили успешностью проводимых противоэпидемических мероприятий. В средине XIX столетия чуму отнесли к категории вымерших болезней , карантины закрыли за ненадобностью.
Однако сапронозная теория поддержания Y.pestis в природе не объясняет ни востребованность и крайне злокачественное течение чумы у людей в отдельные исторические эпохи, ни предопределенность в пандемической смене патогенов, сопровождавших пандемии чумы, ни появление легочных осложнений после столетий господства бубонной чумы.
Многокомпонентный пандемический процесс
В 1436 г. знаменитый врач Конкорегио, выживший в период масштабных чумных и оспенных эпидемий, заметил, что эпидемия оспы нередко оказывается предвестницей эпидемии чумы и, что чума в таких случаях оказывается более опасной. Среди врачей этого столетия бытовало твердое убеждение, что variola (оспа) и morbilli (корь) могут встречаться одиночными случаями в любое время, но когда эти болезни распространяются весьма сильно, то в этом случае они являются предвестниками большой эпидемии чумы (Губерт В., 1896).
Здесь мы подходим к рассмотрению очень интересной закономерности - не только вторая, но и первая и даже так называемая третья пандемии чумы происходят на одном и том же фоне пандемического распространения совершенно разных, и вроде бы никак не связанных между собой, контагиозных болезней.
Первая пандемия
Ей предшествуют пандемии проказы и натуральной оспы. К началу первой пандемии чумы ( Чума Юстиниана , 533-588), проказа представляла серьезную проблему для европейцев, об этом в частности, говорят свидетельства о принимаемых властями законах, требующих отделения прокаженных от здоровых людей. Аналогичную ситуации мы наблюдаем и с заболеваемостью натуральной оспой. Только начиная с первых десятилетий VI-го столетия можно констатировать фактическое существование оспы в Европе (Губерт В., 1896).
В начале VII столетия масштабные эпидемии чумы в Европе прекратились (Гезер Г., 1867). В VIII-Х столетиях сообщения об оспенных эпидемиях в Европе исключительно редки (Губерт В., 1896). Судя по тому, что среди историков долгое время бытовало убеждение в том, что проказа проникла в Европу только во время крестовых походов (первый состоялся в 1096 г.), то можно утверждать, что с VIII по начало XI столетия она переживала период упадка (Гезер Г., 1867).
Вторая пандемия
Ей вновь предшествуют те же пандемические события, что и Юстиниановой чуме. Начиная с XI столетия, в Европе вновь активизируется проказа. В конце XII столетия, по неизвестным причинам, после почти двухсотлетней спячки , начали расти контагиозность и вирулентность возбудителя оспы, что нашло отражение в летописях (Гезер Г., 1867). Черная смерть начинается на фоне масштабных оспенных эпидемий, свирепствующих по всему Европейскому континенту.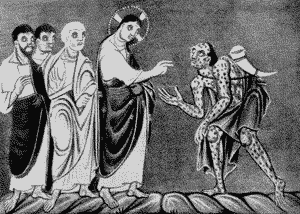
Иисус Христос с учениками и прокаженным. Прокаженный с рогом перед Христом. Из немецкой рукописи, подготовленной до 1000 г. Пандемия «черной смерти» совпала с небывалой по злокачественности пандемией проказы
Третья пандемия
В средине XIX столетия оспа, чума и проказа считались побежденными болезнями. Падение заболеваемости натуральной оспой в начале XIX столетия достоверно зафиксировано в странах, где тщательно велся статистический учет причин смертности. Например, в Лондоне смертность от оспы стала уменьшаться еще с 70-х гг. XVIII века (Duncan S.R. и Scott S., 1996), т.е. до введения в практику метода вакцинации, предложенного Эдвардом Дженнером только в 1796 г. Падение заболеваемости оспой в Швеции началось еще в 80-х гг. XVIII столетия. К 1800 г. заболеваемость снизилась почти в 2,5 раза, но тогда в Швеции не было еще ни одного вакцинированного ребенка (Бразоль Л., 1875).
Исчезновение же проказы началось в конце XV столетия, прежде всего, в Италии, несколько позже во Франции, потом в Голландии и Северной Германии. В 1664 г. во Франции, по распоряжению Людовика XIV, был закрыт последний лепрозорий за отсутствием больных (Гезер Г., 1867).
Накануне третьей пандемии чумы (вторая половина XIX столетия) стала повсеместно возвращаться проказа (Тржецяк С., 1913). Оспенная пандемия неожиданно возобновилась в 1871 г. Эта катастрофа произошла на фоне принятых в европейских странах законах об обязательной вакцинации детей и проведении повторной вакцинации взрослых. Болезнь свирепствовала два года, приняв такой злокачественный характер, который не наблюдался даже в XVIII веке (Бразоль Л.Е., 1875).
Приведенные данные свидетельствуют о возможности многокомпонентных пандемических процессов, проявляющихся периодическим и сменяющим друг друга популяционным повышением восприимчивости населения к контагиозным болезням двух типов: с длительным инкубационным периодом (например, для проказы он составляет 3-7 лет, для туберкулеза до трех месяцев) и относительно коротким инкубационным периодом (натуральная оспа, 8-14 суток; SARS, 4-8 суток; грипп в форме испанки , 1-4 суток и др.). Причем процесс пандемической активации контагиозных болезней инерционен и специфичен. Неконтагиозная, бубонная форма чумы, появляется на этом фоне как востребованная случайность. Синхронность масштабного появления чумы и проказы, объясняется тем, что возбудители этих болезней являются природно-очаговыми сапронозами. Следовательно, та же последовательность событий, которая приводит к колебательным процессам в экосистемах лепрозного микроба, оказывает аналогичное действие и на экосистему простейшие-Y.pestis .
Одновременно можно предположить участие на пандемическом поле VI и XIV столетий еще каких-то других игроков , дополнительных к известной эпидемической триаде природный резервуар возбудителя чумы - его переносчики - восприимчивая человеческая популяция . Собирательно назовем их фактором Х .
Фактор Х в пандемиях чумы
Благодаря этому фактору особое преимущество в эпидемических цепочках приобретают исключительно высоковирулентные штаммы возбудителя болезни, либо они становятся высоковирулентными из-за повышенной специфической восприимчивости человеческих популяций, либо клиника болезни настолько меняется, что начинает восприниматься как совсем другая болезнь .
Легочные осложнения бубонной чумы. Рассмотрим несколько крупных вспышек бубонной чумы с легочными осложнениями, описания которых достаточно информативны.
Вспышка чумы в Ветлянке
Легочная чума - болезнь отдельных генотипов. По клиническим проявлениям болезни эпидемия чумы в Ветлянке делится на два периода: 1) с преобладанием бубонных форм (с 28 сентября); 2) с преобладанием безбубонных и, в частности, легочных форм (Минх Г.Н., 1898).
Второй период эпидемии начался в многочисленной семье богатого казака, Осипа Белова. Эпидемия проявила себя сразу массой почти внезапных смертельных заболеваний с почти полным отсутствием бубонов. Минх констатировал тот факт, что близкое соседство даже с наиболее пострадавшими от чумы семействами, не имело никакого влияния на распространение болезни по станице. Ему также удалось установить, что основной принцип ее движения - не соседские или внутридомовые отношения между заболевшими, а родственные. В 1878 г. Минх не знал таких терминов как генотип, поэтому он сделал вывод, что легочная чума распространяется по родословным . Следовательно, второй период ветлянской чумы начался со случайного вовлечения в эпидемический процесс особых человеческих генотипов, у которых инфекционный процесс проявился вторично-легочной формой болезни. После достижения критической массы из вторично-легочных случаев болезни, чума сформировала самостоятельные эпидемические цепочки, состоящие уже из первично-легочных случаев.
Незаразная легочная чума в Анзобе.
Вспыхнула в 1897 г. в горном кишлаке Анзоб (Таджикистан) находящемся в непосредственной близости от открытого только в 1970-х гг. Гиссарского природного очага чумы. По подсчетам Я.М. Финкельштейна (1906), из 400 человек населения кишлака, чумой заболело 250 жителей (62%), из них выжили только 13. По окончанию эпидемии общая смертность составила 85%. Путем опроса родственников погибших он установил, что из 204 (93%) заболевших легочной чумой (кашель, кровохарканье, отсутствие бубонов), не выжил никто (смертность 100%); из 14 анзобцев, заболевших бубонной чумой, выжили 8 (смертность 43%); один житель, у которого отмечали и бубоны и кровохарканье, погиб.
Другое обстоятельство, обратившее тогда на себя внимание Финкельштейна (1906), это не заразность больных с вторично-легочной чумой. Это касалось как русского медицинского персонала, так и здоровых анзобцев, согласившиеся ухаживать за больными днем, но на ночь уходившие в свои семьи. Аналогичные наблюдения сделаны во время эпидемий легочной чумы в Пали в 1836 г. (см. выше) и в Колобовке в 1899 г. (Розанов П.Г., 1900).
Таким образом, возвращение легочной чумы происходит на территориях реликтовых очагов чумы в населенных пунктах, где население представлено родственниками, или имеет относительно однообразный генофонд. Внешне такая вспышка выглядит семейной , однако она имеет очень упорное течение и свои клинические особенности, зависящие от генотипического состава пораженной чумой популяции. В основе распространенности феномена спонтанного возникновения вторично-легочной чумы во время эпидемий бубонной чумы, лежит популяционная частота встречаемости генов, экспрессия которых в ответ на инфекцию сопровождается воспалительными некрозами легочной ткани. В подтверждение этого своего предположение, нам представляется весьма полезным привести данные из работы Geiger K. и Sarvetnic N. (1996). В опытах на трансгенных мышах они показали, что сверхэспрессия IL-2 сопровождается поражением ЦНС и развитием воспалительных процессов (пневмоний) даже без дополнительного инфицирования их возбудителем какой-либо болезни (!). Известно, что у человека бактериемия появляется уж на ранней стадии развития бубонной чумы и в этот процесс, как правило, вовлекаются бронхолегочные лимфатические узлы. Утрата же некротизированными участками легочной ткани способности сопротивляться возбудителю болезни, значительно облегчает развитие в ней специфических очагов поражения.
Количество и патологическая анатомия таких участков в легких, а также заразность заболевшего чумой человека зависят от экспрессии дополнительных аллелей генов, продукты которых в повышенных количествах причастны к деструкции легочной ткани. Например, во время бубонной чумы среди китайцев в Гонконге в 1896 г. поражений легких практически не было. На 240 вскрытий обнаружено 5 случаев воспалительной инфильтрации в нижних долях легкого (Вильм, 1898). Во время бубонной чумы среди малороссов во Владимировке в 1900 г. (Волго-Уральский природный очаг чумы), осложнения со стороны легких были весьма распространены (3 на 6 вскрытий). В легочной ткани находили уплотнения круглой формы, величиной от лесного ореха до куриного яйца, и даже почти всей нижней доли. Но ни в одном случае у больных не наблюдали кровохаркания. Однако почти одновременно вспыхнувшая на территории этого же природного очага чума среди казахов Таловского округа Внутренней Киргизской Орды, имела исключительно легочный характер (Страхович И. В., Гос В.И., 1907).
Следовательно, основное осложнение бубонной чумы во времена черной смерти - вторично-легочная чума, стала следствием инерционного генетического процесса (один из составляющих фактора Х ), приведшего к преобладанию в популяциях населения Европейского континента носителей аллелей генов, экспрессия которых в ответ на инфекцию сопровождалась воспалительными некрозами легочной ткани (предположительно, IL-2). За это говорит и то обстоятельство, что во время первых вспышек черной смерти погибали, в основном, молодые люди, носители соответствующих аллелей генов. В 1918-1920 гг. аналогичная возрастная структура заболевших наблюдалась во время пандемии испанки . Однако расшифровка генома этого вируса, выполненная во второй половине 1990-х гг., не позволила установить мутации, которые могли бы объяснить смертоносный характер этой пандемии (Reid et al., 2002).
Вне эпидемического процесса такие мутации можно считать нейтральными. В Европе и России их носители были элиминированы Y.pestis уже в первой половине XV столетия, после чего чума (до Ветлянской эпидемии, 1878) проявлялась только в бубонной и бубонно-септической формах, какой бы масштаб она не принимала.
Интоксикация при чуме
Уже более столетия, начиная с работ Минха (1881, 1898) и до учебника Шуваловой Е.П. с соавт. (2001) включительно, авторы-клиницисты настаивают на том, что в основе патогенеза чумы лежит выработка ее возбудителем каких то токсинов. Однако бактериологи до настоящего времени так и не нашли у Y.pestis истинных экзотоксинов (какими, например, являются дифтерийный или столбнячный), способные в опытах на животных вызывать основные симптомы, наблюдаемые при чуме. Твердое следование догме, что именно токсин или какое-то другое вещество, образуемое Y.pestis, определяет патологический процесс, привело к тому, что в исследовании механизмов патогенеза при чуме не было сделано никаких принципиальных открытий. Как проявление кризиса изучения специфических механизмов интоксикации при чуме Домарадский И.В. (1998) рассматривает смещение интереса ученых с такого специфического токсина, каким является мышиный , к неспецифическому фактору патогенности любой грамотрицательной бактерии - липополисахаридному комплексу бактериальной стенки. Тем самым он признает, что так называемая интоксикация при чуме носит неспецифический характер.
Действительно, многие симптомы в клинической картине чумы наблюдаются при других инфекционных болезнях, в том числе тех, возбудителей которых никак не заподозришь в синтезе истинных экзотоксинов (например, вирусы).
Рассмотрим такой важный синдром клиники чумы, как сердечно-сосудистый, обычно относимый к проявлениям интоксикации . Быстро нарастающее поражение сердечно-сосудистой системы, резкое падение кровяного давления, помрачение сознания и геморрагии ( чумные знаки ) у некоторых больных чумой развиваются даже раньше, чем бубоны или осложнения со стороны легких. Но они встречаются и при большинстве вирусных инфекций, например, при гриппе (особенно во время пандемии испанки ), и хантавирусных инфекциях; и при паразитарном заболевании - малярии. В то же время эти неспецифические симптомы проявляются далеко не у всех больных, и в разные эпидемии их частота варьирует (Шувалова Е.П. с соавт., 2001). Следовательно, их проявление все же зависит от каких-то специфических факторов, которые могут проявляться как в клинике болезни, так и в ее эпидемиологии.
В поисках ответа на вопрос, что это за специфический фактор ( фактор Х ), от которого зависят неспецифические клинические проявления болезни, обратимся к аналогиям.
Лица, которые экспрессировали гены главного комплекса гистосовместимости В8(HLA-B8), переносили хантавирусную инфекцию более тяжело, чем экспрессировавшие HLA-B27. У них болезнь проявлялась пониженным кровяным давлением, повышенным содержанием креатинина в крови. Одновременно наблюдалось большее количество вируса в моче и крови, определяемого с помощью полимеразной цепной реакции (ПЦР). У лиц с HLA-B27, болезнь протекала в мягкой форме. Почти все больные с гаплотипом HLA-A1-B8-DR3, у которых прогрессивно развивались все те же симптомы шока (наблюдаемые и при чуме; см. работу Шуваловой с соавт., 2001), экспрессировали вторую аллель фактора некроза опухолей (TNF2) (Wilson A. еt al.,1997).
От распространенной аллели TNF1, TNF2 отличает более сильный транскрипционный активатор, способствующий увеличению экспрессии фактора некроза опухолей (TNF-?). Патологическим проявлением повышенной экспрессии TNF-? становится более тяжелое течение малярии и хантавирусного синдрома. Причем при обеих болезнях наблюдаются неспецифические симптомы : нарастающее поражение сердечно-сосудистой системы, резкое падение кровяного давления, помрачение сознания, геморрагии и быстро наступающая смерть. Малярия, как правило, приобретает церебральную форму (McNicholl J. et al., 1997). Можно утверждать, что знаки при чуме представляют собой неспецифическую реакцию, зависящую от наличия в геноме человека дополнительных копий специфической аллели (TNF2 и др.), имеющих различную встречаемость и соотношение в человеческих популяциях в различные исторические эпохи.
Проведенный нами анализ описаний эпидемий чумы конца XVIII - начала ХХ столетий также показал, что характерное для черной смерти обильное появление на коже больных чумных знаков (петехий, бубонов, карбункулов), из-за которых труп умершего человека выглядел черным, не находится в зависимости от легочных осложнений (Самойлович Д., 1952; Вилле Я.П., 1828; Гезер Г., 1867; Минх Г.Н., 1898; Заболотный Д.К., 1899; Дербек Ф.И., 1905; Богуцкий В.М., 1911; и др.). Следовательно, эти типы осложнений течения бубонной чумы зависят от аллелей различных групп генов.
Востребованность чумы
Заключается в наличии в восприимчивой популяции особей, подбор возбудителей контагиозных инфекций для которых осуществляется специфическим образом, за счет структур фактора Х . Эпидемический процесс сначала запускается по неспецифическому механизму, т.е., в результате каких-то случайных обстоятельств возбудитель болезни проникает в группу восприимчивых особей, но затем его уже поддерживает фактор Х . Отсюда крайне упорное пандемическое течение черной смерти и оспы в средние века и туберкулеза в настоящее время.
Обратимся к хорошо изученным аналогиям среди современных пандемий. Обычно исследователи обращают внимание на последовательность вторжения в человеческий организм возбудителей СПИД-ассоциированных инфекций (их не менее 18) и даже устанавливают некоторую зависимость от количества в крови человека Т-лимфоцитов. Однако их удивляет то обстоятельство, что в самом перечне таких инфекций нет никакой логики. Возбудители крайне таксономически разнородны, у них отсутствуют видимые признаки сходства в жизненных циклах и экологии (Лысенко А.Я. с соавт., 1996).
А между тем, перечень все же информативен. По нему, по крайней мере, можно судить о том, что таксономия, жизненный цикл и экология этих возбудителей не являются предопределяющими в развитии СПИДа. Следовательно, существуют специфические причины, следствием действия которых именно эти паразиты становятся востребованными ВИЧ-инфицированной составляющей человечества.
ВИЧ, как паразитический организм, должен постоянно разрушать клетки хозяина и потреблять высвобождающуюся биохимическую энергию. Для этого он индуцирует в инфицированных макрофагах, клетках микроглии и астроцитах синтез большого количества цитокинов (факторы некроза опухолей, интерлейкины и др.). В норме их действие на уровне центральных органов иммунитета (костный мозг, тимус) является точечным и импульсным. При ВИЧ-инфекции оно становится непрерывным, постоянно оказывая токсическое действие на организм человека. Но одновременно цитокины (IL2 и IL3) являются сильными факторами роста Leismania major (СПИД-индикаторная инфекция) - благодаря этому она получает преимущества в размножении перед другими таксономически сходными организмами (Mazingue C. et al., 1989), т.е. оба цитокина выступают для лейшманий в качестве фактора Х , поэтому распространение лейшманиоза приобретает параллельный ВИЧ, специфический пандемический характер.
Для возбудителя другой СПИД-индикаторной инфекции - M. avium, известны, по крайней мере, два фактора Х . Первый, это интерлейкин 6 (IL6). Повышенный синтез этого лимфокина у ВИЧ-инфицированных людей резко увеличивает чувствительность макрофагов к M. avium (Crowle A.J. et al., 1991). Второй - белок gp120 самого ВИЧ. При сравнительных исследованиях бронхоальвеолярного смыва инфицированных и здоровых лиц, было установлено, что присутствие этого белка усиливает размножение M.avium в альвеолярных макрофагах (Denis M., 1994).
Роль одного из факторов Х для пандемического распространения возбудителя туберкулеза играет сурфактантный белок A (SP-A), присутствующий в бронхоальвеолярной жидкости ВИЧ-инфицированных людей. Он усиливает прикрепление M. tuberculosis к альвеолярным макрофагам (Downing J.P. et al., 1995), по этой причине пандемия туберкулеза, всегда считавшегося социальной болезнью, имеет крайне упорное течение в богатых странах с развитым здравоохранением. Не привлеки в начале 1970 гг. ретровирусы внимание онкологов, современная инфицированность населения туберкулезом рассматривалась бы учеными как самостоятельное явление, правда, очень злокачественное.
Между чумой и оспой в отношении взаимодействия их возбудителей с общими рецепторами на поверхности клеток-мишеней может оказаться больше сходства, чем между возбудителями токсоплазмоза, криптоспоридоза и норвежской чесотки, распространяющихся параллельно СПИДу. Не исключено, что фактор Х сам может индуцировать образование рецепторов для вируса натуральной оспы и Y. pestis на поверхности клеток макроорганизма, критически важных для инициации инфекционного процесса.

«Человек, умирающий от чумы». Рисунок из коллекции английских религиозных текстов (XV столетие), собранных в Йоркшире монахами ордена картезианцев (одеяние картезианца — длинное белое платье с белым капюшоном, а вне монастыря — черное). Священник выполняет последний обряд над телом человека, умершего от чумы в монастыре говоря: «Прими тело в могилу, спаси Христос твою душу». В это же время смерть пронзает человека в постели своим копьем: «Я сражаюсь с тобой много дней для того, что бы ты стал моей добычей». Произведение принадлежит Британской библиотеке, Лондон.
Современные представления о кожных болезнях позволяют утверждать, что за больных проказой в Средние века могли приниматься больные СПИДом с лимфомами, проявлениями кожного туберкулеза, с пневмоцистозом, с кандидозом и гистоплазмозом, т.е. болезнями, считающимися сегодня СПИД-индикаторными . Наличие некоторых мутаций (ССR5 и др.) в геноме человека белой расы свидетельствуют о том, что в природе должны существовать какие-то очень древние терминаторы пандемий СПИДа и подобных болезней. Ими как раз и могут быть те контагиозные инфекции, которые оставили после себя столь дурную славу в прошлом - натуральная оспа, распространяющиеся пандемически чума и туберкулез (Супотницкий М.В., 2000).
Однако то, что СПИД или какая-то СПИД-индикаторная инфекция были проявлением фактора Х , который специфически спровоцировал распространение проказы, натуральной оспы, и черной смерти, пока только осторожная гипотеза, подтверждение или опровержение которой возможно в рамках палеовирусологических и палеобактериологических методов исследования.
Была ли третья пандемия чумы ?
Если считать третью пандемию явлением хронологически самостоятельным, то тогда между ней и второй пандемией чумы должен быть временной промежуток, который можно охарактеризовать как прекращение чумы . Новая пандемия по масштабу охваченных ею территорий должна, по крайней мере, превышать масштабы активизации очагов чумы последнего всплеска их активности в прошлую пандемию. Такую границу можно провести между первой и второй пандемиями, но вот с третьей это сделать невозможно. И вот почему.
Развитие первой и второй пандемий на Европейском континенте напоминает лесной пожар, который разгорелся на огромной территории в течение 5 лет. Даже уже два этих дискретных временных явления заставляют предположить наличие каких-то очень сложно связанных друг с другом природных явлений, которые, достигнув некой критической массы в течение неизвестного периода времени (годы, десятилетия, столетия?) приводят к многолетней глобальной активизации реликтовых очагов чумы. По территориальному распространению вторая пандемия, начавшись в средине 1340-х гг. в Индии и Китае, то прекращаясь, то начинаясь вновь, но уже на значительно больших территориях, достигла своего максимума к 60-м гг. XIV столетия.
Дальнейшее развитие второй пандемии больше соответствует сложно периодическим колебаниям в еще неизвестных экокосистемах, вмещающих чумной микроб, чем передаче возбудителя болезни по цепочке от человека к человеку или из крысиных очагов чумы посредством инфицированных эктопаразитов. На огромных территориях вспыхивают синхронизированные по времени, но разные по интенсивности эпидемии чумы. Между ними остаются временные промежутки, которые достаточны для того, что бы сменилось несколько поколений людей, живших в мире без чумы . Например, относительно благополучными по чуме для Европы были промежутки между 80-ми гг. XIV столетия и 40-ми гг. XV столетия; между 1527 г. и 1545 г. На территории отдельных реликтовых очагов чумы можно отметить и более продолжительные периоды эпидемического благополучия. До чудовищной чумы 1773 г. в Багдаде и Басре, жители Месопотамии не встречались с ней почти 90 лет. Между последними чумными эпидемиями в Москве (1654 и 1771 гг.) прошло 117 лет, за это время сменилось 4 поколения москвичей, не знавших этой болезни. Пульсация реликтовых очагов чумы возобновляется на огромных территориях - например в средине XVII этот процесс осуществился на пространстве от Вятки до Лондона в течение 10 лет, погубив, по приблизительным оценкам, до 1 млн. человек. Всего же таких периодов (пиков активности чумы) после окончания черной смерти 1346-1351 гг. и последовавших за ней пульсаций очагов чумы в более северных широтах (первый пик - 1346-1382 гг.), можно насчитать еще не менее трех: 1440-1530 гг.; 1545-1683 гг. (по площади охваченных чумой территорий его можно прировнять к черной смерти); 1710-1830 гг. Внутрь этих пиков всеевропейских пульсаций вложены пики пульсаций отдельных релитиктовых очагов. Однако, со средины XVII столетия, постепенно происходит отступление чумы в направлении с севера и запада на юго-восток. Все больше западных и северо-западных территорий Европы становятся свободными от чумы , если, конечно, под чумой понимать только ее вспышки среди людей.
Угасая на Европейском континенте, очаги чумы разгораются южнее, в Индии и Китае. Эти территории были интенсивно поражены чумой во время черной смерти, но в XVII веке Индостан и Китай оставались относительно благополучными по чуме. В начале XIX столетия, когда, разбросав свои смертельные искры, угасли балканские и причерноморские очаги чумы, чума в Индии вновь резко активизировалась. В эти же годы началась активизация природных очагов чумы на юге Китая и в Индокитае. К 1834-35 гг. относятся сведения о появившейся в китайском городе Нинпо (Нинбо - город на берегу Восточно-Китайского моря) эпидемии чумоподобного характера . В 1850 г., когда бубонная чума выпала из поля зрения европейских ученых, и, даже, считалась ими вымершей болезнью , легочная чума вновь вспыхнула на южных склонах Гималаев (в Гурвале и Камауне). В этот же год бубонная чума напомнила о себе в Кантоне (Китай). В 1858 г. обнаружилось движение чумы на север. Она появилась в бубонной форме среди бедуинов на прибрежье Средиземного моря в близи Триполи, чем вызвала замешательство в стане победителей чумы . Еще через 5 лет активизировались очаги Великого Евразийского чумного излома (1863-1879), с начала 1880-х гг. чума все чаще регистрируется в Северном Прикаспии, Поволжье, Монголии, Северном Китае и Забайкалье. Со средины XIX столетия чума становится эндемичной на юге Китая. В 1864 г. вновь чумоподобная эпидемия в Нинпо. В 1866 г. чума обнаружена в столице китайской провинции Юннань (граничит с Вьетнамом и Бирмой) - в городе Юннаньфу. Во время гражданских войн в 1860-70 гг. чума свирепствовала на юге Китая эндемически. В 1867 г. чума вспыхивает в Пакхое (город в провинции Юннань на берегу Тонкинского залива). В 1871 г. чума возвращается в провинцию Юннань, значительные вспышки в Пакхое. С конца 70-х годов XIX столетия обнаружилась сезонность в появлении чумы в южном Китае. Она стала аккуратно появляться в январе каждого года в провинции Квангси и в округах Лиенчоу и Лейчоу. Болезнь наблюдалась ежегодно с 1881 г. по 1884 г. в Пакхое. В 1889 г. была эпидемия в Люнгчоу (Лунчжоу), а в 1890 г. чума вспыхнула в Ву-чу (Бучу), на берегу между Пакхоем и Кантоном. В 1891 г. появилась чума в Као-чау, области смежной с Лиенчоу, где находится Пакхой. В 1892 г. вспыхнула эпидемии чумы в Ан-пу, городе, находящемся в 180 км к востоку от Пакхоя. В 1893 г. вновь появилась чума в Пакхое. Она также была в Юннане, преимущественно в городе Мингз (в южной части провинции). Чума свирепствовала в Люнгчоу и во многих городах провинции Квангси. В 1894 г. чума устраивает побоище в китайских портовых городах Кантоне, Гонконге и Амое, и только тогда (!) она попадает в поле зрения европейских ученых, как начавшаяся третья пандемия . С этим событием по времени совпало бактериологическое обнаружение Y.pestis у крыс в городах, охваченных чумой.
За первые 10 лет пандемии (1894-1904) чума отметилась в 87 (из нескольких тысяч!) портовых городов. Хотя в те же годы фиксировали эпидемии, которые не были связанны с морскими портами, все равно действовал определенный стереотип мышления: пандемия чумы началась в 1894 г. и была портовой (Ахшарумов Д.Д., 1900; Диаптроптов П.И., 1901; Wu Lien-Ten et al., 1936; Николаев Н.И., 1949). Факты, свидетельствовавшие о том, что чума в XIX столетии не прекращалась, не принимались во внимание, так как они противоречили ставшей очень прогрессивной теории разноса чумы кораблями, кстати, известной еще со времен чумы в Марселе 588 г. (см. у Гезера Г., 1867). Следовательно, началом третьей пандемии чумы сегодня считается случайно взятая дата одной из эпидемий чумы в Кантоне, периодически вспыхивающих там с 1850-х гг.
Сопоставление же территориальных масштабов так называемой третьей пандемии с предыдущими пиками активности чумы второй пандемии , показывает, что она значительно уступает по этому показателю последнему, четвертому ее пику (1710-1830). Учитывая также и искусственность даты начала третьей пандемии , правильнее будет считать ее не самостоятельным явлением, а пятым, самым низким пиком второй пандемии чумы. Более важным признаком для проведения разграничительной линии между пандемиями, отражающим серьезное изменение генотипического состава человеческих популяций, мы считаем качественное изменение эпидемического процесса при чуме - возвращение легочной чумы в начале XIX столетия в Индии.
В 1346-1351 гг. население Европы было инфицировано возбудителем чумы из активизировавшихся на ее территории природных очагов чумы, существующих неопределенно долго и вне зависимости от популяций диких и домашних грызунов, и их эктопаразитов. Такие очаги (предлагается называть их реликтовыми), возможно, и по сегодняшний день сохраняют Y.pestis в некультивируемом состоянии в одноклеточных почвенных животных. Черная смерть - это патологический синдром, возникающий во время вспышек бубонной чумы у людей с определенными генотипами. Он характеризуется развитием вторичной чумной пневмонии, геморрагическим проявлениями, симптомами шока и обширными кожными поражениями (карбункулы, петехии). Эпидемическая выраженность черной смерти зависит от количества людей, носителей определенных аллелей генов, проживающих на территориях пульсирующих очагов чумы. Продолжительность активизации реликтовых очагов чумы во время ее второй пандемии составляла 400-500 лет. Существуют неизученные факторы, делающие востребованными возбудителей контагиозных инфекций отдельными человеческими популяциями (предлагается объединить их под общим названием фактор Х ). Во время каждой из известных пандемий чумы они проявляли себя в рамках многокомпонентного пандемического процесса. Начало третьей пандемии чумы можно отнести на второе десятилетие XIX столетия, когда произошло качественное изменение клиники чумы, вызванное накоплением определенных генотипов среди людей - болезнь стала проявляться во вторично-легочной форме.
Литература
Авиценна (Абу Али Ибн Сина). Канон врачебной науки. Книга IV, часть первая. - Ташкент, 1980.
Анисимов А.П. Факторы Yersinia pestis, обеспечивающие циркуляцию и сохранение возбудителя чумы в экосистемах природных сообществ. Сообщения 1 и 2 // Молекулярная генетика, микробиология и вирусология. - 2002. - № 3, С. 3-23; № 4, С. 3-11.
Архангельский Г.Ф. Амбулантные формы чумы (pestis ambulans) и её значение в эпидемиологии //Сборник сочинений по судебной медицине, судебной психиатрии, медицинской полиции, общественной гигиене, эпидемиологии, медицинской географии и медицинской статистике. - 1879. - Т. 1. - С. 132-193.
Ахшарумов Д.Д. Чума последних годов XIX столетия. - Полтава, 1900.
Богуцкий В.М. Эпидемия чумы в Харбине и его окрестностях. Харбин, 1911.
Балтазар М. (Baltazard M.) Стойкость чумы в постоянных очагах // Журн. гиг. эпидемиол. (Прага). - 1964. - Vol. 8, P. 333-343.
Боккаччо Дж. Декамерон. - Кишинев, 1982.
Борисенков Е.П., Пасецкий В.М. Тысячелетняя летопись необычайных явлений природы. - М, 1988.
Бразоль Л.Е. Дженнеризм и Пастеризм, критический очерк оснований оспопрививания. - Харьков, 1875.
Васильев К.Г., Сегал А.Е. История эпидемий в России. - М., 1960.
Вильм. Чумная эпидемия в Гонконге в 1896 году // Медицинское прибавление к морскому сборнику. - 1898. - Сентябрь. - С. 254 - 266; Ноябрь. - С. 319 - 327.
Вилле Я.П. Практические замечания о чуме. 2-е издание. М.: 1828.
Гезер Г. История повальных болезней. - Санкт Петербург, 1867.
Гирш А. Индийская чума // Военно-медицинский журнал. - 1853. - Часть LXII (III). - С. 29-48.
Губерт В.О. Оспа и оспопрививание. - Санкт Петербург, 1896.
Дербек Ф.И. История чумных эпидемий в России. - Санкт Петербург, 1905.
Диаптроптов П.И. Обзор заболеваний чумой на побережье Средиземного моря и в портовых городах Европы в 1900 и в начале 1901 гг. //Русский архив патол., клин., мед. и бактериол. - 1901 . - Т. 12. - С. 77-94.
Домарадский И.В. Чума. - М., 1998.
Дятлов А.И. Эволюционные аспекты в природной очаговости чумы. - Ставрополь, 1989.
Заболотный Д.К. Пустулезная форма чумы //Русский архив патологии. - 1899. - Т. VIII. - С. 239-242.
Козлов М.П., Султанов Г.В. Эпидемические проявления чумы в прошлом и настоящем. - Махачкала, 1993.
Лысенко А.Я., Турьянов М.Х., Лавадовская М.В. и др. ВИЧ-инфекция и СПИД-ассоциируемые заболевания. - М., 1996.
Маренникова С.С., Щелкунов С.Н. Патогенные для человека ортопоксвирусы. М.,1998.
Минх Г.Н. Отчет об Астраханской эпидемии. - М., 1881.
Минх Г.Н. Чума в России (Ветлянская эпидемия 1878-1879 гг.). - Киев, 1898.
Миронов Н.П. О существовании в прошлом природной очаговости чумы в Южной Европе // Журн. микробиол., эпидемиол. и иммунобиол. - 1958. - № 8. - С. 18-23.
Николаев Н.И. Чума в Манчжурии. Дис... д-ра мед. наук.- Киров, 1949.
Никульшин С.В., Онацкая Т.Т., Луканина Л.М. Изучение ассоциаций почвенных амеб H.rhisodes с бактериями - возбудителями чумы и псевдотуберкулеза в эксперименте // Журн. микробиол.- 1993. - № 9-10.- С. 2-5.
Петтенкофер М. Холера.- СПб., 1885.
Рихтер А. История медицины в России.- М., 1814.
Руднев Г.П. Клиника чумы.- М., 1940.
Розанов П.Г. Отдельное мнение о Колобовской эпидемии // Больничная газета С.П. Боткина.- 1900, 12 июня.
Самойлович Д. Рассуждения о чуме, производившей в 1771 г. опустошения в Российской империи и особенно в столичном городе Москве // Избранные произведения. - М., 1952. - Т. 2. - С. 8-278.
Страхович И. В., Гос В.И. Чумные эпидемии в Астраханской губернии в 1900-1901 гг. //Сборник работ по чуме. Вып. 2. - 1907. - СПб. - С. 43-92.
Сунцов В.В., Сунцова Н.И. Экологические аспекты эволюции микроба чумы Y. рestis и генезис природных очагов // Общая биология. - 2000. - № 6. - С. 645-657.
Супотницкий М.В. Микроорганизмы, токсины и эпидемии.- М., 2000.
Тржецяк С. Проказа в библейские времена и в настоящее время, с обращением особого внимания на положение вопроса о проказе в России (Доклад, читанный в заседании I Биологического Отделения Рус. Общ. охр. нар. здравия 6 апреля 1913 г.). - СПб., 1913.
Федоров В.Н. К вопросу о существовании в прошлом в Европе природной очаговости чумы // Гигиена, эпидемиология, микробиология и иммунология (Прага).- 1960.- Вып. 4, С. 132-137.
Финкельштейн Я.М. Еще раз о чуме в Анзобе // Военно медицинский журнал. - 1906. - Т. 217, № 9. - С. 27-57.
Шувалова Е.П., Белозеров Е.С., Беляева Т.В. и др. Инфекционные болезни. - Ростов на Дону, 2001.
Щепотьев Н.К. Чумные эпидемии в России. - СПб., 1897.
Шпилевский С.М. О чуме в Казани в царствование Алексея Михайловича // Сборник статей о чуме, изданных Обществом Казанских врачей. Казань, 1879. - С. 53-55.
Cole S.T., Buchrieser С. // Nature. - 2001, Vol. 413. - Р. 467-469.
Crowle A.J., Cochn D.L., Poche P. Defects in sera from AIDS patients and from non-AIDS patients with Mycobacterium avium infection which decreases macrophage resistance to M.avium // Infect. Immun. - 1991. - Vol. 57, № 7. - P. 1445 - 1451.
Denis M. Envelope glycoprotein (gp120) from HIV-1 enhances Mycobacterium avium growth in human bronchoalveolar macrophages // Clin. and Exp. Immunol. - 1994. - Vol. 98, ? 1. - P. 123-127.
Duncan S.R., Scott S. Oscillatory dynamics of smallpox and the impact of vaccination // J. Theor. Biol.- 1996. - Vol.183. - P.447-454.
Geiger K., Sarvetnic N. The Influence of Cytokines on the Central Nervous System of Transgenic Mice // Transgenic Models of Human Viral and Immunological Disease. - California, 1996.
Kortepeter M., Christopher G., Cieslak T. et al. USAMRIID'S Medical Management of Biological Casualities. Handbook. - Fort Detrick. - 2001.
Lederberg J. Infectious diseases as an evolutionary paradigm // Emerging Infectious diseases. - 1997. - Vol. 3, № 4. - P. 1-9.
McNicholl J., Smith D.K., Qari S. et al. Host Genes and: The Role of the Chemokine Receptor Gene CCR5 and Its Allele (32CCR5) // Emerging Infectious diseases. - 1997. - Vol. 3, № 3. - P. 261 - 272.
Mollaret H. Conservation experimentale de la pest dans le sol // Bull. Soc. Phatol. Exot. - 1963. - Vol. 56. - P. 1168-1182.
Reid Ann H., Fanning T.G., Janczewski T.A. et al. Characterization of the 1918 Spanish Influenza Virus Matrix Gene Segment // J. оf Virology. -, 2002 - Vol. 76, № 21. - Р. 10717-10722.
Vogel U., Claus H. The evolution of human pathogens: examples and clinical implications // Int. Med. Microbiol. - 2000. - Vol. 290. - P. 511-518.
Wilson A., Symons J., McDowell T. et. al. Effects of polymorphism in the tumor necrosis factor alpha promoter on transcriptional activation // Proc. Natl. Acad Sci. USA. - 1997. - Vol. 94, №. 10. - P. 3195 - 3199.
Wheelis M. Biological Warfare at the 1346 Siege of Caffa // Emerging Infectious diseases. - 2002. - Vol. 8, № 9. - P. 971-975.
Wu Lien-Ten U.A., Chun J.W., Pollitzen R. et al. Plague. A manual for medical and public health workers. - Shanghai, 1936.
ПРИЛОЖЕНИЕ 1
Современные представления о распространении черной смерти построены только на одном свидетельстве, нотариуса генуэзских работорговцев де-Мюсси, наблюдавшего ее в Кафе (Феодосия) в 1346-1348 гг. Складывается впечатление, что других документов нет. Но это не так. В 1348 г. Парижский медицинский факультет, по повелению короля Филиппа, обнародовал свое мнение о чуме для публичного поучения народа, как следует держать себя относительно болезни . Документ был опубликован французским богословом Ж. Мишоном (1806-1881) по оригиналу, хранящемуся в Парижской императорской библиотеке (Documents inedits sur la grand peste de 1348. Paris, Londres et New-York, 1860). Русский перевод мы приводим по тексту Г.Ф. Архангельского (1879).
Мнение членов Парижского медицинского факультета о происхождении эпидемии чумы черной смерти и о предохранительных мерах против нее
Мы, члены Парижской медицинской коллеги, по зрелом обсуждении и глубоком рассмотрении теперешней смертности, и согласно с мнением наших древних учителей, полагаем обнародовать причины этого чумного мора (pestilence), по законам и принципам астрологии и естественных наук. Вследствие сего, мы заявляем следующее: известно, что в Индии и в странах великого моря, небесные светила, которые борются с лучами солнца и с жаром небесных огней, оказывают специально их влияние на это море и сильно борются с его водами. От того рождаются испарения, которые помрачают солнце и изменяют его свет в тьму. Эти испарения возобновляют свое поднятие и свое падение в течение 28 дней непрерывно; но, наконец, солнце и огонь действуют так сильно на море, что они вытягивают из него большую часть вод и превращают эти воды в испарения, которые поднимаются в воздух, и если это происходит в странах, где воды испорчены мертвыми рыбами, то такая гнилая вода не может быть ни поглощена теплотою солнца, ни превратиться в здоровую воду, град, снег или иней; эти испарения, разлитые в воздухе, покрывают туманом многие страны. Подобное обстоятельство случилось в Аравии, в части Индии, в равнинах и долинах Македонии, в Албании, Венгрии, Сицилии и Сардинии, где ни одного человека не осталось в живых; то же самое будет во всех землях, на которые будет дуть воздух, зачумленный Индийским морем, пока солнце будет находиться в знаке льва. Если жители не будут соблюдать следующие предписания или другие аналогичные, то мы возвещаем им неизбежную смерть: если только милосердие Христа не призовет их к жизни какие-либо другим образом.
Мы думаем, что небесные светила, вспомоществуемые природой, делают усилия, в своем небесном могуществе, для покровительствования человеческому роду и для исцеления его болезней и, вместе с солнцем, для проникания силою огня, чрез густоту тумана в продолжение десяти дней и до 17-го числа ближайшего месяца июля. Этот туман превратится в гнилой дождь, падение которого очистить воздух; тотчас как гром или град возвестить его, каждый должен остерегаться этого дождя, зажигая костры из виноградных ветвей, лаврового или другого зеленого дерева; равно пусть жгут в больших количествах полынь и ромашку на общественных площадях и в местах многолюдных; пусть никто не выходить в поле прежде, нежели совершенно не высохнет земля и 3 дня после того, каждый в это время пусть позаботится принимать немного пищи и остерегаться утренней, вечерней и ночной прохлады. Пусть не едят ни живности, ни водяных птиц, ни молодой свинины, ни старого быка, в особенности же жирного мяса. Пусть употребляют мясо животных, одаренных натурой горячей и сухой, но не горячащей, ни раздражающей
Мы рекомендуем приправы с толченым перцем, корицу и пряности, особенно лицам, которые привыкли ужинать немного и из отборных блюд; спать днем вредно; пусть сон продолжается только до восхода солнца или немножко позже. Пусть мало пьют за завтраком, ужинают в 11 часов и могут во время стола пить немножко больше, чем утром; пусть пьют вино чистое и легкое, смешанное с шестою частью воды; фрукты сухие и свежие, употребляемые с вином, не вредны, без вина же они могут быть опасны. Красная морковь и другие овощи, свежие или маринованные, могут быть вредны; растения ароматические, такие как шалфей и розмарин, напротив здоровы; съестные припасы холодные, водянистые или влажные вообще вредны. Опасно выходить ночью и до 3-х часов утра по причине росы. Не должно есть никакой рыбы, излишнее упражнение может повредить; одеваться тепло, остерегаться холода, сырости, дождя, ничего не варить на дождевой воде, принимать за столом не много териака; оливковое масле в пище смертельно; тучные люди пусть выходят на солнце; очень большое воздержание, беспокойство духа, гнев и пьянство опасны; дизентерии должно бояться; ванны вредны; пусть поддерживают желудок свободным при помощи клистиров; сношение с женщинами смертельно. Эти предписание применимы особенно для тех, которые живут на берегах моря или на островах, на которые подул гибельный ветер.
Уже из самого содержания документа видно, что Парижский факультет излагал свое мнение о чуме черной смерти еще до ее появления во Франции, и именно в то время, когда она опустошала Турцию, Венгрию, Италию и Сардинию. Даже это обстоятельство свидетельствует о том, что завоз чумы галиотами из Кафы, не более чем поздняя историческая мистификация, удобная для рассмотрения причин появления чумы в Европе с позиций контагионистического учения. В 1347 г. власти Франции знали о начавшейся пандемии и готовились ей противодействовать.
Факультет заявил и то, что он высказывает свое мнение, согласуясь с мнением древних своих учителей , а между тем, местом родины появившейся черной смерти он обозначает не Эфиопию или Левант, как считали эти учителя , а Индию и страны великого моря (видимо, ими так назывался Тихий и Индийский океаны), т.е. они твердо знали регион, где началось это бедствие. Среди ученых господствовала миазматическая теория, объяснявшая причину возникновения эпидемий гнилостью воздуха и поступательным движением зачумленного воздуха Индийского моря на материк Азии и Европы. До приобретения практического опыта борьбы с чумой, она не считалась контагиозной болезнью. Появление чумы в Китае определяется 1346 г., следовательно, в один и тот же год она появилась в Европе (в Крыму, Сицилии, Прикаспии, на Северном Кавказе), Индии, Закавказье и в Малой Азии, т.е., вторая пандемия началась как глобальная катастрофа.
ПРИЛОЖЕНИЕ 2 (готовится)
Предназначалась для журнала «РЭТ-инфо», но не была в нем опубликована
Вышла наша книга Супотницкий М.В., Супотницкая Н.С. Очерки истории чумы: В 2-х кн. Кн. I : 468 с.: ил.; Кн. II : 696 с.: ил.; М., 2006.
На сайте выложены очерки:
V. «ЧЕРНАЯ СМЕРТЬ» — ВТОРОЕ ПРИШЕСТВИЕ ЧУМЫ В ЕВРОПУ (1346—1351)
X. ЧУМА В МАРСЕЛЕ И ПРОВАНСЕ (1720—1722)
XX. ТРАГЕДИЯ В СТАНИЦЕ ВЕТЛЯНСКОЙ — ТУПИК ДОБАКТЕРИАЛЬНЫХ ПРЕДСТАВЛЕНИЙ ОБ ЭПИДЕМИОЛОГИИ ЧУМЫ (1878)
XXX. ЛАБОРАТОРНАЯ ЧУМА В ФОРТЕ «АЛЕКСАНДР I» (1904—1907)
XXXI. ЭПИДЕМИЯ ЛЕГОЧНОЙ ЧУМЫ В МАНЬЧЖУРИИ И ЗАБАЙКАЛЬЕ (1910—1911)
XXXIII. ЗАБЫТЫЕ ЭПИДЕМИИ ЛЕГОЧНОЙ ЧУМЫ НА ДАЛЬНЕМ ВОСТОКЕ (1921—1922)
XXXIV. ЧУМА ОТ ДЬЯВОЛА В КИТАЕ (1933—1945)
XXXV. ЭПИДЕМИИ ЧУМЫ В МАНЬЧЖУРИИ В 1945—1947 ГГ. — ПЕРЕЛОМ В ЛЕЧЕНИИ И В ПРОФИЛАКТИКЕ ЧУМЫ

Михаил Васильевич Супотницкий
Российский микробиолог, полковник медицинской службы запаса, изобретатель, автор книг и статей по истории эпидемий чумы и других особо опасных инфекций, истории разработки и применения химического и биологического оружия. Заместитель главного редактора научно-практического журнала «Вестник войск РХБ защиты» Министерства обороны РФ.