Историческая информация о вспышках чумы как источник идей для будущих научных открытий в чумологии

«Мысль о присутствии чумы очень неприятна с непривычки …»
А. С. Пушкин, 1835
История эпидемий не преподается в медицинских ВУЗах, да и учеными она нередко воспринимается просто как интересное чтение. Между тем на фоне уже сложившегося «мелкотемья» в изучении эпидемических процессов, сведения, содержащиеся в исторических описаниях эпидемий, могут серьезно повысить мотивацию молодых исследователей, дать им возможность обосновать и спланировать темы НИР, способные приблизить их к научным открытиям.
Методология исследования
В исторических источниках выявлялись те сведения о развитии эпидемических и инфекционных процессов, которые не могут быть объяснены с точки зрения представлений об этих процессах, распространенных в современных учебниках и монографиях по инфекционным болезням и эпидемиологии. Так же выявлялись несовпадения в описаниях одних и тех же явлений в самих исторических источниках. Объяснение таких «нестыковок» давались на основе представлений, взятых из других областей знания, имеющих хорошие методические заделы и позволяющие осуществлять экспериментальную проверку этих предположений и гипотез; а также путем аналогий с другими, хорошо изученными объектами. Под «историческими источниками» в данной работе понимаются труды ученых и врачей либо самостоятельно описавших отдельные эпидемии чумы, либо выполнивших их обобщающие описания по трудам других очевидцев эпидемических событий.
Глобальная цикличность эпидемий чумы
Историческими источниками глобальная цикличность эпидемий чумы зафиксирована как пандемии: первая — чума Юстиниана (531–589), вторая — «черная смерть» (1346–1351) и третья — «портовая чума» (1894–1904). Их суть заключалась в том, что на огромных территориях планеты в сравнительно короткий исторический период вспыхивали эпидемии чумы, сопровождающиеся чудовищной смертностью населения отдельных городов и, даже, стран. Причины их возникновения объяснялись, как это не покажется сегодня странным, сходным образом на протяжении почти 1400 лет — чума распространялась кораблями. Различия состояли лишь в деталях. Считалось, что чума в Марсель во время первой пандемии была занесена неким кораблем (Гезер Г., 1867). Вторая пандемия объясняется злонамеренными действиями татарского хана Джаныбека, который применил якобы бактериологическое оружие (БО) против жителей города Каффы (нынешняя Феодосия). После чего заболевшие легочной чумой жители разнесли ее на судах по всей Европе. Хотя исторические источники свидетельствуют о развитии в европейских городах преимущественно бубонной чумы, версия о применении БО в средине XIV в. сегодня подается как абсолютно достоверная (см. в работах Онищенко Г. Г. с соавт., 2003; Черкасского Б. Л., 2002; Wheelis V., 2002). Третья пандемия объясняется с позиций представлений начала ХХ столетия о роли грызунов как природного резервуара возбудителя чумы, но корабли, как основной фактор распространения чумы, по-прежнему в версии остаются. Больные чумой крысы проникали на торговые суда и ими распространялись по миру. Покинув корабль в том или ином порту, они вызывали чуму у местных крыс, затем посредством инфицированных блох болезнь «перекидывалась» на человека. Одновременно формировался устойчивый природный очаг чумы среди местных грызунов (Федоров В. Н., Козакевич В. П., 1959). Если мы более внимательно посмотрим на эти события в историческом аспекте, то выявляются «выпадения» имеющихся фактов из этих «научных» объяснений, заставляющие все же сомневаться в их правильности.
Я искренне признателен доктору географических наук Дмитрию Васильевичу Николаенко за предоставленную возможность опубликовать свою статью в ежегоднике "Энвайронментальная эпидемиология и медицинская география".
Во-первых, эпидемии чумы развиваются тысячелетиями на одних и тех же местностях. Так первая и вторая пандемии чумы вспыхивают в одних и тех же городах, примерно в одной и той же территориальной последовательности (с юга на север и с востока на запад) и охватывают Европейский континент за 3–5 лет (см., например, работу Гезера Г., 1867). Даже так называемая «третья пандемия», не достигшая масштабов двух предыдущих, по своим контурам с ними все же совпадает. Наряду с этим многие авторы свидетельствуют о наличие местностей, куда чума «никогда не заносится» (Гирш А., 1853; Архангельский Г. Ф., 1879; Щепотьев Н. К., 1884, 1897).
Работая над монографией «Очерки истории чумы», мы не обнаружили ни одного исторического свидетельства, бесспорно подтверждающего, что чума в Европе начиналась именно в портах. Если изучать детали появления чумы в том или ином портовом городе, то как правило, территории портов меньше всего поражались чумой. Например, в Одессе в 1812 г. бубонная чума началась среди актеров театра (при полном благополучии по чуме в одесском чумном карантине!), но еще раньше она началась в отдаленных от Одессы местностях Северного Причерноморья и Малороссии (Белиловский В. А. с соавт., 1904). Тем не менее, во всех источниках ее называют «портовой». Во время чумы в Глазго в 1900 г. в городе и в порту не было найдено ни одной чумной крысы, ни свидетельств ее «заноса» больными чумой людьми (Диаптроптов П. И., 1901).
Весьма интересно то обстоятельство, что возбудитель чумы (Yersinia pestis) может сохраняться на одних и тех же территориях без его поддержания среди грызунов. И.В. Домарадский (1998) описал «пробуждение» одного из известных очагов чумы в районе Гулженги в Забайкалье, на протяжении 13 лет никак не обозначавшего себя эпизоотиями среди грызунов.
Во-вторых, пандемии чумы не начинаются внезапно, они имеют длительный «период предвестников», когда исторические хроники за несколько десятилетий до объявленной пандемии фиксируют локальные вспышки чумы в отдельных населенных пунктах, не связанные с портами. В Средние века их приписывали колдовству и магии злонамеренных лиц. Сами «злонамеренные лица» в подвалах инквизиции быстро признавали свою вину. Таким образом, причины и этих вспышек чумы были очевидны для современников, и эпидемиология чумы находилась в самодостаточном состоянии (о том как добивались признательных показаний от «сеятелей чумы» см. в работах Киттериджа Дж. Л., 2005; и Трахтенберга Дж., 1998).
В третьих, все пандемии чумы проявляются множеством несвязанных между собой вспышек бубонной чумы. По наблюдениям современников, чума обычно «двигалась скачками». С одного населенного пункта в другой, нередко пропуская другие, густонаселенные, лежащие на пути ее «перемещения». Даже развивающиеся в некоторых городах вспышки легочной чумы, как это, например, имело место во времена «черной смерти», все же происходили на фоне масштабных вспышек бубонной чумы и имели четкие территориальные и временные рамки (Гезер Г., 1867; Гирш А., 1853; Вогралик Г. Ф., 1935).
Бубонная чума не контагиозна и никогда не выходит из своих природных очагов. Но природные очаги, наличие которых могло бы объяснить почти 500-летнее ее присутствие на Европейском континенте, сегодня встречаются только на юге Европейской России. Западная, Центральная и Восточная Европа, наиболее пострадавшие от эпидемий чумы в XIV–XVIII вв., не имеют ее природных очагов, по крайне мере таких, в которых бы возбудитель чумы поддерживался грызунами.
Обращает внимание и то обстоятельство, что известные сегодня природные очаги чумы территориально занимают значительно меньшие площади (их современные границы см. в фундаментальной монографии Козлова М.П., Султанова Г.В., 1993), чем те, границы которые можно очертить по историческим хроникам. Например, в первой половине XIX в. Приараксинский равнинно-предгорный очаг чумы проявлял себя бубонными случаями болезни до верховий рек Карс-чай и Аракс-чай на Карском плато. А Закавказский равнинно-предгорный очаг распространялся от Апшеронского полуострова до предгорий Сурамского хребта (описание вспышек чумы на этих территориях см. в работе Дербека Г.Ф., 1905).
И последнее (четвертое) «выпадение» имеющихся фактов из «научных» объяснений эпидемиологии чумы — масштабные эпидемии чумы развиваются на фоне вспышек других опасных болезней. Специалистам по чуме известно, что для возникновения даже единичного бубонного случая болезни среди людей, должны совпасть редко встречающиеся в природе обстоятельства (наличие в природном очаге восприимчивых к чуме грызунов, развитие чумы у грызуна в септической форме, блохи должны быть «блокированы» и др.). Поэтому большая часть вспышек чумы ограничивается небольшим количеством вовлеченных в нее людей (десятки, и очень редко, сотни случаев болезни). Анализ же исторических источников показывает, что эти маловероятные события масштабируются только при наступлении других эпидемических событий, совпадения с которым нет смысла игнорировать.
Первая и вторая пандемии чумы развивались на фоне масштабных эпидемий проказы, или той болезни, которая тогда считалась проказой. Другим пандемическим событием — предвестником чумы, средневековые авторы считали распространение среди людей натуральной оспы. В 1436 г. знаменитый врач Конкорегио (J. M. Concoregio, 1405–1448), выживший в период масштабных чумных и оспенных эпидемий, заметил, что эпидемия оспы нередко оказывается предвестницей эпидемии чумы и, что чума в таких случаях оказывается более опасной. Среди врачей бытовало твердое убеждение, что variola (оспа) и morbilli (корь) могут встречаться одиночными случаями в любое время, но когда эти болезни распространяются весьма сильно, то в этом случае они являются предвестниками большой эпидемии чумы (Губерт В., 1896).
Примерно за два века до пандемии чумы в исторических источниках появляются упоминания о распространении проказы. Правительствами принимаются соответствующие законы, открываются тысячи лепрозориев в странах, где сегодня о существовании проказы даже не подозревают. За несколько десятилетий «до чумы» активизируется натуральная оспа и только на фоне этих двух бедствий, появляется третье — чума в ее пандемическом распространении (см. хронологию у Гезера Г., 1867).
Накануне «возвращения» чумы во второй половине XIX столетия она, как и оспа, считались уже уничтоженными болезнями. Победа над чумой приписывалась карантинам, и их повсеместно закрыли за ненадобностью; исчезновение натуральной оспы объяснялось вакцинацией. Например, в 1867 г. В. Веревкин писал: «Читая в настоящее время о тех громадных потерях, какие терпело человечество от этих ужасных болезней, невольно задаешь себе вопрос: куда же они подевались? От чего они теперь не с такою же силою свирепствуют в народе? Конечно, ответа на эти вопросы мы должны искать, между прочим, в успехах цивилизации современного человечества». В те годы спорной была заразность проказы. Многие ученые склонялись к тому, что эта болезнь передается по каким-то наследственным механизмам, так как ее эпидемические цепочки среди людей не прослеживались (см. в кн. Васильева К.Г., Сегала А.Е., 1960). Однако глобальная эпидемическая ситуация неожиданно поменялась уже к концу 1860-х гг.
Жестокая пандемия натуральной оспы 1870–1874 гг. напомнила современникам оспенные пандемии средневековья, вот только разница между ними была в том, что эта пандемия развилась уже на фоне массовых противооспенных вакцинаций и ревакцинаций населения (Бразоль Л. Е., 1875). С конца 1870-х гг. активизировалась чума (эпидемии в Южном Китае, на севере Персии, в Астраханской губернии, в Маньчжурии и др. местах). Вновь «проснулась» проказа. После почти 200-летнего перерыва она появилась в Прибалтике. Значительно увеличилось количество выявленных больных проказой в Астраханской губернии, на Кавказе и в Закавказье (Васильев К. Г., Сегал А. Е., 1960).
Даже эти исторические сведения позволяют предположить существование в природе совсем иных очагов чумы, чем известные сегодня в рамках учения о природной очаговости чумы Д. К. Заболотного. Такие очаги существуют тысячелетиями вне связи с наличием на данных территориях инфицированных возбудителем чумы грызунов. Они проявляют себя периодически возникающими на одних и тех же территориях масштабными эпидемиями чумы. Активизация таких очагов зависит от неизвестных пока факторов, но эти же факторы влияют и на активизацию неизвестных пока природных очагов возбудителей натуральной оспы и проказы. Для того, что бы отграничить их от очагов чумы, выявляемых бактериологическими методами по эпизоотиям чумы среди грызунов, я предлагаю назвать их реликтовым. К уточнению этого термина мы вернемся ниже, а пока попытаемся очертить границы реликтовых очагов по упоминаниям эпидемий чумы в исторических источниках, не вдаваясь в объяснение того, каким образом на этих территориях может поддерживаться Y. pestis.
Реликтовые очаги чумы
По историческим источникам нам удалось проследить такие территории, начиная с пандемии «черной смерти» (см. Супотницкий М.В., 2003, 2005; Супотницкий М.В. и Супотницкая Н.С., 2006). Их максимально возможные границы выглядят следующим образом.
В России…
Реликтовый Северо-западный природный очаг чумы — представляет собой восточную оконечность цепочки очагов чумы, тянущихся от побережья северной Франции (пролив Ла-Манш), через территории Голландии, Дании, германских государств, балтийское побережье Швеции и Польши, Прибалтику и по Карелии до южного побережья Белого моря. Активизация очага зафиксирована в русских летописях XIII в. и приходится на начало малого ледникового периода. В 1424 г., судя по территории, охваченной чумой, пульсация очага достигла максимума — бубонная и легочная чума в Новгороде, Пскове, Карелии, а также в «Литве и Немцах». Последняя эпидемия чумы в Пскове пришлась на 1711 г., в Новгородской области на 1606 г.
Реликтовый природный очаг чумы Русской равнины. Включает территорию, по форме приближающуюся к четырехугольнику с углами Великие Луки—Вологда—Кострома—Смоленск. Вместе с похолоданием климата постепенно разогревался очаг чумы. В 1229—1230 гг. чума дала о себе знать мором в Смоленске, где погибло до 32 тыс. человек. «Черная смерть» 1346–1351 гг. обошла Москву. В 1363 г. чума поразила Переславль, Коломну, Владимир, Суздаль, Дмитров, Можайск, Вологду. Только в 1364 г. чума появилась в Москве: «Того же лета бысть мор велик во граде Москве и по всем волостям московским (Никон. лет.)». В этом же году чума свирепствовала в Переславле, Рязани, Коломне, Суздале, Дмитрове, Можайске, на Волоке «и во все грады разыдеся мор силен и страшен… и бысть пустыни вокруг непроходимыа (Никон. лет.)». В 1654–1656 гг. пульсация очага достигла максимум. В Москве эпидемия началась в августе 1654 г. и продолжалась до января 1655 г. погубив до 300 тыс. человек. С августа по сентябрь 1654 г. чумой были охвачены Кострома, Тверь, Коломна, Торжок, Звенигород, Кашин, Переславль-Рязанский, Переславль-Залесский, Суздаль, Тула, Калуга, Углич, многие их слободы и уезды. Осенью 1771 г. в Москве и в ближайших к ней городах разгорается эпидемия чумы, сопровождающаяся бунтом населения. Однако по охвату территории ее масштабы были значительно меньшими, чем в 1654 г. В 1773 г. пульсация очага прекратилась.
Реликтовые природные очаги чумы Северного Причерноморья и Малоросии. Это восточная оконечность последовательности реликтовых очагов чумы, тянущихся от Балкан на восток вдоль всего северного побережья Черного и Азовского морей до очагов чумы Великого Евразийского чумного «излома» (см. ниже). На севере границы очага можно расположить на равнинном пространстве между Волыно-Подольской и Среднерусской возвышенностями до границ Курской области. Пульсации очагов происходят в направлении с запада на восток. Начало активизации очагов чумы Северного Причерноморья и Малоросии приходится на период свирепствования «черной смерти». В 1738 г. их активность достигла максимума. Во второй половине XVIII в. чума постепенно отступает в направлении с севера на юг и дробится на мелкие очажки. В первой половине XIX в. активными остаются очаги чумы Северного Причерноморья, расположенные на пространстве между Крымским полуостровом и Тирасполем, причем их границы постепенно сужаются. Небольшую активность эти очаги проявили в начале ХХ в. (чума в Одессе в 1902 и 1910 гг.)
Великий Евразийский чумной «излом». Под ним мы понимаем гигантскую последовательность как реликтовых, так и пульсирующих сегодня природных очагов чумы, расположенную «изломанной» дугой — от пустынь Йемена до возвышенностей, называемых Северными увалами (север Кировской области). Эта последовательность включает Саудовско-Йеменский очаг с центром в Эль-Асире и Хавлане, Сирийско-Месопотамский пустынный очаг, Курдо-Иранский природный очаг чумы, очаги чумы Центрально-Иранского плоскогорья, Северо-Иранских краевых гор, а также смежных с ними плоскогорий и впадин Афганистана, Приараксинский участок очаговости чумы, Закавказский равнинно-предгорный очаг чумы, Закавказский высокогорный природный очаг чумы, Центрально-Кавказский природный очаг чумы, Восточно-Кавказский природный очаг чумы, Прикаспийский северо-западный очаг чумы, Зауральский природный очаг чумы, Волго-Уральский природный очаг чумы и реликтовые очаги чумы долин рек Ветлуга, Вятка и Кама до Северных Увалов. Объединение в «излом» не носит произвольный характер. В пульсациях этих очагов в максимумы их активности (XVII–XIX вв.) отчетливо прослеживается пространственная и временная последовательность.
Активизация очагов «излома» начинается с горного Курдистана. Затем, в промежуток времени в 2–4 года, в этот процесс вовлекаются Сирийско-Месопотамские очаги чумы, поэтому в восприятии современников чума идет с верховьев Тигра и Евфрата по их долинам и опустошает Месопотамию, юго-западную Персию и Малоазиатскую часть Турции. В промежуток времени 10–15 лет от начала активизации очагов горного Курдистана, чума «поднимается» на север, вспыхивая сначала на Карском плато; затем на территориях, которые сегодня называют Приараксинским участком очаговости чумы; затем, в течение еще двух-трех десятилетий, она «скачками» продвигается далее на север; и, описав дугу вокруг Каспия со стороны Кавказского хребта, «ударяет» по Астрахани (1662, 1693, 1727, 1806, 1877), после чего поднимается по Волге, иногда до Саратова (1808), реже, в максимум активности «излома», ещё дальше (Нижний Новгород, 1363; Казань, 1665; Вятка, 1667).
Современники «черной смерти» зафиксировали активизацию, по крайней мере, части очагов чумы Великого Евразийского чумного «излома» — от Курдо-Иранского до Прикаспийского Северо-западного. В их восприятии это была «другая волна чумной эпидемии», прокатившаяся через Сирию, Армению, Малую Азию и прикаспийские земли.
На территории России наиболее северным участком «Излома», на котором в 1346 г. наблюдалась пульсации реликтовых очагов чумы, видимо, была местность в районе Енотаевки (Астраханская область), где по свидетельству летописца произошла «казнь от Бога» на жителей города Бездеж (Воскр. лет.). Самой южной границей «Излома» можно считать местность между реками Тигр и Евфрат в районе Багдада, где в 1347 г. вспышки бубонной и бубонно-септической чумы ужаснули население повальной смертностью.
В годы последующих пульсаций реликтовых очагов «излома», территории пульсирующих очагов расширялись и смещались на север и на юг. В 1363 г. их границы «поднялись» до Нижнего Новгорода: «…а пришел (мор) от низу, от Бездежа, …и опусте земля вся и порасте лесом, и бысть пустыни всюду непроходимы (Никоновская летопись)». До средины XVII в. о чуме на территории «излома» в исторических источниках не упоминается, возможно, это не от невнимательности летописцев. Например, Петр Петерей (1620) писал: «Московиты, находящиеся за Рязанью и в Татарии, вовсе не знают моровой язвы, страдают же от нее лишь близкие к западной границе, а именно: Новгород, Псков, Смоленск и др.» т. е. в этот период активными оставались очаги чумы, расположенные в северной части Европейского континента.
Пульсации очагов «излома» возобновились в средине XVI в. Их максимум на территории России совпал с наибольшей активностью реликтового природного очага чумы Русской равнины и большинства других европейских реликтовых очагов. Летом 1655 г. моровое поветрие появилось в низовьях Волги и в Астрахани. Затем эпидемия вспыхнула Казани, где в тот год от чумы погибло 48 тыс. человек. В 1657 г. чума свирепствовала и в соседней с Казанской — Вятской области (северная граница «излома»), и в самом городе Вятке. Летом 1657 г. чума снова возобновилась в низовьях Волги, в 1692 г. чума повторилась в Астрахани.
Южная граница реликтовых очагов «излома» проходит по побережьям Персидского залива (на восточном побережье, по крайней мере, она достигает Бушера). Максимум в их активности достигнут через столетие после максимума активности северных очагов «излома». Он проявился чудовищными эпидемиями чумы в Багдаде и Басре в 70-х гг. XVIII в. (до 2-х млн. погибших). В последующие годы «северные территории» пульсирующих очагов «излома» (Россия), сужались и «смещались» к югу (сегодня это Прикаспийские, Волго-Уральские и Кавказские очаги чумы), «южные» (Месопотамия), наоборот, «подтягивались» на север (Закавказские и Иранские очаги чумы). Количеств жертв чумы в одних и тех же природных очагах снижалось от вспышки к вспышке, как и их территориальные масштабы.
В Европе…
Балкано-Придунайские реликтовые очаги чумы. Синхронно активизировались как в первую, так и во вторую пандемии чумы, приводя к массовой гибели людей. Среди них можно выделить три группы реликтовых очагов: 1) цепочка очагов на равнинной местности между реками Прутом и Серетом, расположенных от Хотина до Измаила (включая Яссы, Браилов, Галац и др.), своей восточной частью вплотную прилегающих к реликтовым очагам Северного Причерноморья и Малороссии; 2) цепочка очагов на равнинной местности между Дунаем и Черноморским побережьем от Бабадага до Варны (включая Черноводы, Каварну, Кюстенджи); 3) отдельные очаги на равнинной местности между Константинополем, Андрианополем и побережьем Мраморного моря. Максимум активности достигнут в конце XVIII в. Очаги третьей группы проявляли небольшую активность даже в начале ХХ в.
Реликтовые очаги чумы долины реки По. Исторически они наиболее смертоносные в Европе и самые активные на Апеннинском полуострове; многократно разгорались в первую и вторую пандемии. Их пульсациями объясняются сокрушительные эпидемии Средневековья и эпохи Возрождения в городах Венеция, Милан, Падуя, Болонья, Пьяченца, Палермо, Верона и др., иногда воспринимаемых современниками как «конец света». Максимум активности достигнут в средине XVII в.
Реликтовые очаги чумы долины реки Арио. Катастрофические эпидемии чумы в Пизе и Флоренции во времена первой и второй пандемий. Пик активности приходится на времена «черной смерти» (художественное описание см. у Дж. Боккаччо «Декамерон»).
Реликтовые очаги чумы долины реки Вольтурно. Их пульсации проявлялись тяжелыми эпидемиями в Неаполе в период второй пандемии чумы. Самая смертоносная эпидемия чумы вспыхнула в 1656 г. Она унесла жизни 200 тыс. человек из 400 тыс. населения города. Последний раз очаги пульсировали в 1900 г.
Реликтовые очаги чумы долины реки Рона. По многочисленным историческим источникам очень активны. Пульсации проявлялись сокрушительными эпидемиями чумы во времена первой и второй пандемий. Границы очагов можно заключить в треугольник «Авиньон (вершина) — Тулон и Монпелье (основание)». Максимум активности пришелся на 1720–1722 гг.
Сицилийские реликтовые очаги чумы. Располагаются вдоль северного побережья острова между городами Трапани и Сиракузы. Наиболее активно пульсация очагов происходила в период второй пандемии на местности, прилегающей к городу Мессина.
Реликтовые очаги чумы Пиринейского полуострова. Долина реки Дуэро (местность в районе города Порто) — в последний раз пульсировали в начале ХХ столетия; долина реки Гвадалквивир (Севилья); местность южнее и восточней Андалузких гор, включающая Гибралтар, Малагу, Альмерию; низовья реки Турия (Валенсия); долина реки Эбро (Сарагоса); местность, прилегающая к Каталонским горам с востока (Барселона) — в последний раз пульсировали в конце августа 1931 г., когда было выявлено 5 случаев бубонной чумы (двое заболевших умерли) среди тряпичников поселка Госпиталет в 4 км на восток от Барселоны. Пик активности реликтовых очагов чумы Пиринейского полуострова приходится на средину XVII в.
Реликтовые очаги чумы долины реки Гаронна. Пульсации проявлялись сокрушительными эпидемиями чумы в городах Бордо и Тулуза во времена второй пандемии. Пик активности приходится на конец XVII в.
Реликтовые очаги чумы северо-восточной Франции. Это обширное равнинное пространство между городами Руан, Амьен, Реймс и Париж, включающее долины рек Сена, Марна, Сомма. Мощные пульсации во времена первой и второй пандемий (пик активности приходится на средину XVII в.). В начале 1930-х гг. отдельные случаи бубонной чумы в предместье Парижа Сент Уэн. Объяснить их появление в рамках крысиной теории тогда не смогли.
Реликтовые очаги чумы полуострова Корнуолс (о. Великобритания). Равнинная местность, по форме напоминающая неправильный параллелограмм, расположенная между Бристолем и Уэтмутом, и вдоль юго-восточного склона Корнийских гор, где в августе 1348 г. вымерло почти все население («люди ложились, подобно колосьям под серпом жнеца»).
Реликтовые очаги чумы Юго-Восточной Англии. Расположены на местности, включающей Лондон и его пригороды в радиусе 20 миль от Сити. Пульсации достигли максимума в средине XVII в. В 1900 г. отмечены чумные эпизоотии среди лондонских крыс, объясненные в соответствии с представлениями того времени «заносом чумы кораблями».
Реликтовые центрально-европейские очаги чумы. Цепочка реликтовых очагов, расположенных вдоль северных отрогов Альп (сокрушительные эпидемии чумы второй пандемии в городах Женева, Базель, Берн, Цюрих, Мюнхен, Линц, Вена, Краков), и соединяющихся с западной оконечностью Балканских реликтовых очагов чумы (пик активности приходится на первую половину XVII в.).
Реликтовые очаги чумы долин рек Рейн, Везер, Эльба. Во время второй пандемии их эпидемическая активность нарастала по мере приближения к побережью Балтийского моря (сокрушительные эпидемии чумы второй пандемии в Страсбурге, Франкфурте, Кельне, Ганновере, Гамбурге, Магдебурге и др.). Видимо «сливаются» с цепочкой реликтовых очагов чумы, расположенных вдоль побережий пролива Ла-Манш, Северного и Балтийского морей. Пик активности приходится на первую половину XVII в. Наиболее упорно в Центральной Европе эпидемии чумы «держались» на местности, включающей в себя города Дрезден (1680), Магдебург (1681), Галле (1682), Гальберштад, Брауншвейг, Эрфрут, Нордгаузен и Мюльгаузен (1682–1683) — «Германское чумное пятно». На его территории отдельные вспышки и случаи чумы встречались до 1711 г., после чего она «совершенно исчезла» из Германии.
Если нанести территории этих очагов на географическую карту, то в основном они будут располагаться на низменных территориях, расположенных по долинам рек и вблизи морского побережья. Точные границы реликтовых очагов чумы можно определить с помощью методов молекулярной диагностики, если, конечно, такую задачу перед собой поставить. Теперь попытаемся понять, каким образом на этих территориях может поддерживаться Y. pestis.
Первичный резервуар возбудителя чумы
С конца XIX в. большинством ученых считается, что возбудитель чумы поддерживается в природе грызунами как их облигатный паразит. Но у ученых, основывавшихся на наблюдениях длинных межэпизоотических периодов в появлении чумы (см. выше), появились подозрения о том, что в жизненном цикле Y. pestis должен существовать период, когда этот микроорганизм не поддерживается среди грызунов. Ими были выдвинуты гипотезы, объясняющие существование Y. pestis в форме так называемого фазового паразитизма. В соответствии с ними возбудитель чумы может существовать и как паразит (в организме теплокровного хозяина), и как сапрофит в почве (подробный анализ этих гипотез приведен в обстоятельной монографии Сунцова В. В. и Сунцовой Н. И., 2006). Но тогда ставится под сомнение принцип непрерывности эпизоотической циркуляции возбудителя чумы по схеме «грызун-блоха-грызун», догмат в эпидемиологии чумы, существующий с начала ХХ в. Для современных чумологов покушение на него — это примерно тоже самое, что поставить под сомнение марксистско-ленинскую доктрину в СССР. Поэтому сторонникам фазового паразитизма («сапрофитического подхода», «теллурической чумы») дается обычно дружный и сокрушительный отпор со стороны, по крайней мере, части «чумного сообщества», заключающийся в твердой постановке следующих вопросов:
1) а где доказательства размножения Y. pestis в почве?
2) как может такой полиауксотроф как возбудитель чумы, «не способный синтезировать ряд аминокислот, которые он получает в готовом виде из организма хозяина», существовать вне организма хозяина?
Споры между исследователями такого сложного природного явления как чума, ведутся со времен «черной смерти». Иногда проигравшая сторона оказывается на эшафоте, как, например, судовладелец Шато, обвиненный в 1721 г. властями Марселя в завозе в город чумы на одном из своих кораблей (подробнее см. у Гезера Г., 1867). Важно установить, выигрывает ли от них наука в каждом конкретном случае. Забегая несколько вперед, замечу, что действительно, Y. pestis не может размножаться в почве и не может существовать вне клетки-хозяина. Но и у сторонников схемы «грызун-блоха-грызун» (назовем их «грызуноблохистами») положение двусмысленное. Такая схема была открытием более 100 лет назад. Сегодня работа над ее уточнением, конечно, всегда будет считаться полезным научным занятием, так как это самый простой путь к диссертациям и к статусу «видного ученого». Однако кто же тогда ответит на те вопросы, на которые пытались дать ответ сторонники «сапрофитического подхода»? Многолетние перерывы в эпизоотических процессах в природных очагах чумы существуют в объективной реальности. Будучи никак не объясненными, они подрывают, прежде всего, позиции самих «грызуноблохистов». Судя по данным, взятым мной из исторических источников (см. выше), существуют и другие «загадки чумы», еще более интересные, чем, например, объяснение причин того, почему на данной территории появилась чума среди грызунов, хотя десятки предшествующих лет ее там не фиксировали даже единичными случаями. И мы будем их бесконечно игнорировать ради канонической схемы «грызун-блоха-грызун» с ее несколькими вариантами? Если мы не находим их объяснения в исследованиях чумологов, давайте поищем аналогии среди других, менее задогматизированных объектов исследования, чем чума.
Сегодня бурно развивающимся направлением микробиологии является изучение бактерий в экосистемах, включающих почвенных и водных простейших. Такие простейшие живут в широком интервале условий окружающей среды. Адаптация бактерий к протозойным хищникам происходила миллиарды лет, сами простейшие являются эволюционными предшественниками фагоцитирующих клеток крови многоклеточных организмов (более подробно см. у Купера Э., 1980; и в моей монографии — Супотницкий М. В., 2009). В настоящее время ученые насчитывают не менее четырех форм взаимодействия простейших и микроорганизмов.
Во-первых, отдельные бактерии могут использовать их для увеличения своей численности в окружающей среде, как это показано для Klebsiella aerogenes, паразитирующих в Acanthamoeba castellanii. Во-вторых, некоторые бактерии продуцируют литические компоненты, разрушающие простейших, и тем самым предотвращающие фагоцитоз, например Bacillus liqueniformis синтезирует литический компонент, направленный против Nagleria fowleri. В третьих, между бактериями и простейшими могут устанавливаться эндосимбиотические отношения. Бактерии могут оставаться в простейших длительное время в некультивируемом состоянии. Было установлено экспериментально, что отдельные бактерии могут существовать в простейших в таком состоянии не менее шести лет. В четвертых, в процессе эволюции и коэволюции бактерии могут совершенствовать механизм внутриклеточного существования в амебах и затем, после проникновения в организм позвоночных, пользоваться этим механизмом для выживания в макрофагах (Harb O. et al., 2000; Супотницкий М. В., 2009).
Обзор G. Greub и D. Raoult (2004) включает около 50 видов бактерий и отдельные виды риккетсий и вирусов, способных поддерживаться в простейших. Возбудитель чумы также может существовать в простейших почвы, что показано еще в 1993 г. российскими учеными С. В Никульшиным с соавт. (1993). Тем самым два выше поставленных вопроса снимаются. Y. pestis размножается в почве, но как паразит одноклеточных организмов. Эти организмы компенсируют полиауксотрофность возбудителя чумы. И нет ничего ненаучного в таком объяснении механизмов его поддержании в природе, так как для других опасных для человека микроорганизмов они являются уже давно общеизвестными.
Экспериментальные данные свидетельствуют еще об одном феномене, характерном для паразитизма патогенных для человека микроорганизмов у простейших — упрощении генома паразита при специализации его к своему одноклеточному хозяину. Сравнение геномов Yersinia pseudotuberculosis и Y. pestis для ученого, привыкшего считать, что патогенность микроорганизмов обусловлена приобретением генов факторов патогенности (т. е. усложнением генома), требует преодоления некоторого психологического барьера. Возбудитель чумы, более патогенный для человека и большинства модельных животных, чем возбудитель псевдотуберкулеза, оказывается утратил значительную часть генов, которые традиционно относят к генам вирулентности и патогенности. По данным, обобщенным А. П. Анисимовым (2002), по сравнению с псевдотуберкулезным микробом, Y. pestis утрачивает гены адгезинов, уреазы (сдвиг рамки считывания), инвазинов Inv и Ail (вставка IS-элементов), подвижности, способности к синтезу О-боковых цепей ЛПС (не установленные механизмы образования мутаций) и ряд других. Из 17 «биосинтетических» генов, выявленных у псевдотуберкулезного микроба, 5 в геноме Y. pestis инактивированны за счет вставок и делеций. Компьютерный анализ полного генома чумного микроба (штамм CO92, биовар Orientalis; выделен от человека, погибшего от легочной чумы) показал наличие 149 псевдогенов.
Объяснение этому феномену я вижу в сравнении экологии обоих микробов. У псевдотуберкулезного микроба очень широкий круг хозяев, в основном среди гидробионтов. Он является комменсалом для зоопланктона (дафнии, циклопы), бентосных животных (кольчатые черви, моллюски, личинки насекомых и др.) и высших растений. Возбудитель же чумы, скорее всего, специализирован на узком круге почвенных простейших. Следовательно, упрощение его генома является следствием дегенеративной эволюции, характерной для видовой специализации любого паразита. Но специализации не к отдельным видам позвоночных животных (включая человека), а к простейшим. Для многоклеточных организмов он является случайным паразитом и не поддерживается ими в природе. Т. е. этот микроорганизм является аутохонным компонентом различных экосистем, включающих простейшие, и не нуждается для своего поддержания в природе в циркуляции среди теплокровных организмов.
Выше я писал, что пандемиям чумы предшествуют пандемии проказы и натуральной оспы и что и на возбудитель чумы, и на возбудители этих двух болезней могут влиять сходные факторы, если они находятся в каких-то общих природных резервуарах. Эта гипотеза не выглядит фантастической, если учитывать следующие два обстоятельства.
Первое — для Mycobacterium leprae, микобактерии, вызывающей проказу, было показано, что она может поддерживаться в свободно живущих почвенных амебах. Однако бактериальной репликации или микробного лизиса амеб, вызванного этими микобактериями, описано не было, что свидетельствует в пользу их симбиотических отношений и частично объясняет клинику болезни, вызываемой этим микроорганизмом у людей. Развитие проказы происходит в течение десятков лет, при этом инкубационный период болезни может длиться до пяти лет (Greub G., Didier R., 2004).
Второе — действительно, для вируса натуральной оспы природный резервуар не показан. Вирус считается строгим антропонозоом, к тому же уничтоженным, что можно прочитать в любом учебнике. Но в последние годы выявляются интересные факты по вирусам, близкородственным ортопоксвирусам. В амебах Acanthamoeba polyphaga обнаружен крупный ДНК-вирус (диаметр зрелых частиц достигает 400 нм), названный мимивирусом, т. е. «имитирующий бактерию» (mimivirus — «microbe-mimicking virus»), так как его сначала приняли за бактерию. Таксономически он занимает положение между Poxviridae (к этому таксону относится возбудитель натуральной оспы), Iridoviridae (к ним относятся вирус африканской лихорадки свиней — African swine fever; и многие вирусы земноводных, рыб и насекомых) и Phycodnaviridae (La Scola B. et al., 2003). Но еще более 20 лет назад В. М. Жданов (1988) указывал на то, что ортопоксвирусы по ряду свойств аналогичны бактериям. Это сложно организованные вирусы диаметром частиц 250–390х200–260 нм. Размер их генома сходен с размером генома некоторых бактерий. Репликация ДНК автономна и также практически ничем не отличается от бактериальной. Ортопоксвирусы очень устойчивы во внешней среде.
Если рассматривать ортопоксвирусы в ряду бактерий, то, по мнению В.М. Жданова (1988), последовательность эволюционных событий будет выглядеть следующим образом: риккетсии > хламидии > ортопоксвирусы, т. е. налицо дегенеративная эволюция, когда все более глубокая специализация к конкретному хозяину сопровождается упрощением строения паразита. Для большинства риккетсии внутриклеточный паразитизм обязателен, у хламидий он неизбежен, поскольку они не имеют митохондрий и потому являются энергетическими паразитами. Ортопоксвирусы с их сложной структурой и автономной репликацией ДНК можно представить как дальнейший этап дегенеративной эволюции, сопровождающейся потерей синтезирующего белок аппарата. Для хламидий и риккетсий возможность паразитирования в простейших определенных видов уже показана. В природе Chlamydophila pneumoniae поддерживается в Acanthamoeba castelanii, но не в Parachlamydiaceae или Simkaniaceae (Essig A. et al., 1997). Coxiella burnetii при поглощении почвенными амебами легко в них выживает и размножается (La Scola B., Raoult D., 2001). Описаны также риккетсиоподбные организмы, являющиеся эндосимбионтами Acanthamoeba spp. (Greub G., Didier R., 2004). Поэтому показать облигатный паразитизм возбудителя натуральной оспы у отдельных простейших будет нетрудно, если, конечно, кто-то из ученых этим вопросом займется.
О том, что границы феномена облигатного паразитизма в простейших опасных для человека патогенных микроорганизмов, очень широки, свидетельствуют косвенные признаки. Например, все так называемые возбудители опасных и особоопасных инфекционных болезней (чумы, мелиоидоза, сапа, туляремии, сибирской язвы, бруцеллеза, лихорадки Ку и др.), имеют много сходства в эпидемиологии, биологических свойствах и в клинике вызываемых ими поражений людей. Объяснить их возможно только в том случае, если предположить, что они являются истинными паразитами (или эндосимбионтами) почвенных простейших. В пользу такого предположения говорит «привязанность» вызываемых ими вспышек болезней к конкретным местностям; отсутствие у них резистентности детоксикационного типа к антибиотикам (ненужной для внутриклеточного паразитизма); вовлечение в инфекционный процесс лимфатических узлов и фагоцитирующих клеток крови; а также «запутанность» вопроса о факторах их патогенности и токсинах. Ни один из обнаруженных у них таких «факторов» не может однозначно в эксперименте прояснить клинику вызываемой микроорганизмом у людей болезни, больше похожую на реакцию организма на суперантигенный раздражитель. Не обладают они и контагиозностью, т. е. способностью передаваться от одного теплокровного организма к другому при контакте, что обычно предполагает воздушно-капельный механизм передачи возбудителя болезни непосредственно от одного заболевшего к другому. Редкие случаи «перехода» неконтагиозной бубонной чумы в контагиозную вторично-легочную лишь исключения, подтверждающее выше приведенные наблюдения. Ни больные с такой формой чумы, ни появившиеся через инфицирование от них больные с первично-легочной чумой, не способны поддерживать возбудитель чумы в природе, так как в 100% известных случаев (без лечения антибиотиками) они погибают.
Экспериментальные доказательства способности Y. pestis проникать в организм человека с помощью макрофагов, получены почти 50 лет назад (Cavanaugh D. C., Randall R., 1959). Показано, что продолжительность существования Y. pestis в пределах макрофагов во время инфекционного процесса, зависит от природы хозяина и инфицированных тканей. До 80 % фагосом перитонеальных макрофагов, населенные Y. pestis, сливаются с лизосомами в течение ближайших 2,5 ч после инфицирования, процесс заканчивается в течение 8 ч. Но за это время возбудитель чумы формирует вместительную вакуоль и успевает размножиться (Grabenstein J. et al., 2006). В патогенезе чумной инфекции людей проявляется эволюционно закрепленная за Y. pestis способность к паразитическому существованию среди почвенных простейших, когда микроорганизм не вступает в эндосимбиотические отношения со своим хозяином. Но мир простейших сложен и разнообразен, экологические взаимоотношения между ними изучены плохо. Возможно, что среди одних простейших возбудителю чумы удается существовать как эндосимбионту, среди других, ставших эволюционными предшественниками макрофагов, он размножается как паразит (кстати, взаимоотношения между возбудителем чумы и простейшими разных таксонов — интереснейшая тема для исследований). Но из этого предположения можно объяснить как некоторые особенности чумных эпидемий, так и клиники самой болезни.
Для эпидемий чумы, воспринимаемых современниками как катастрофические (пандемия «черной смерти», вспышки чумы в Лондоне в 1665 г., в Марселе в 1721 г., в Москве в 1771 г. и др.), характерны: 1) постепенное распространение болезни по территории; и 2) цикличность эпидемического процесса. Первые случаи болезни вызываются маловирулентными возбудителями, сотни людей ходят с бубонами, но смертность среди них не высока. Затем вирулентность Y. pestis повышается, смертность среди людей нарастает, бубоны «затвердевают», болезнь проявляет себя в крайне тяжелых формах. По восприятию современников чума на этой стадии эпидемического цикла не поддается лечению, но, достигнув определенного пика, эпидемия резко идет на убыль. Болезнь вновь проявляется в относительно легкой форме. Врачи получают возможность сообщать о своих успехах в лечении чумы с помощью порошка из крабьего глаза, «уксуса четырех разбойников» или просто выжигая бубон раскаленной кочергой.
Первую особенность эпидемий чумы можно объяснить проникновением Y. pestis из экосистем «простейшие–Y. pestis», в которых она существует как эндосимбионт; в простейшие, где она размножается как паразит. Если количество таких простейших в среде, окружающей экосистему «простейшие-Y. pestis», по каким-то причинам достигло неизвестной нам критической массы, то размножившаяся Y. pestis попадает в окружающую среду (в почву). Корнями трубчатых растений бактерия «выносится» на поверхность почвы. Стебли этих растений поедают грызуны и инфицируются Y. pestis. Далее через эктопаразитов инфицировавшихся грызунов возбудитель чумы проникает в дома людей и вызывает среди них вспышку чумы. Ни грызуны, ни их эктопаразиты, не являются первичных резервуаром Y. pestis. Они накапливают возбудитель чумы и приближают его к человеку настолько, что инфицирование людей становится неизбежным.
Вторая особенность эпидемий чумы заключается в постепенном нарастании вирулентности Y. pestis. По аналогии с объяснением повышения вирулентности возбудителя холеры среди гидробионтов, приведенным О. В. Бухариным, В. Ю. Литвиным (1997), этот феномен можно объяснить клональной селекцией вирулентных штаммов Y. pestis среди простейших, в которых она ведет паразитическое существование. Предвидя гневные обвинения «грызуноблохистов» в ненаучности, поясню специально для них, что феномен повышения вирулентности бактерий при их пассировании в простейших уже давно известен. Установлено, что пассирование легионелл в амебах повышает их инвазивность и вирулентность для человека. В пользу последнего предположения говорят экспериментальные данные, показавшие многократно возросшую способность L. рneumophila, выращенных в простейших, внедряться в макрофаги мышей и реплицироваться в легких по сравнению теми же легионеллами, выращенными на искусственной питательной среде (Cirillo J. D. et al., 1994). Также установлено, что M. аvium, выращенная в амебах, более вирулентна чем такая же микобактерия, выращенная на искусственной питательной среде. Причины этого явления следующие. Во-первых, M. аvium, выращенная в амебах, более активно инфицирует как сами амебы, так и клеточные линии интерстинального эпителия (HT-29) и макрофаги. Во-вторых, такие M. аvium обладают расширенными возможностями по колонизации кишечника мышей, в минимальных дозах вызывают у них микобактериальную инфекцию, и быстро проникают в печень и селезенку (Greub G., Didier R., 2004).
Снижение вирулентности Y. pestis во время вспышек чумы в городах, проявляющееся тем, что болезнь начинает появляться в легкой форме, вызвано как повышением резистентности самих свободно живущих простейших, так и грызунов, вовлеченных в процесс передачи этого микроорганизма людям (гибель высокочувствительных к Y. pestis грызунов и простейших, формирование иммунных к возбудителю чумы популяций грызунов и др.). Высоковирулентные штаммы возбудителя чумы сами обрывают цепочки, по которым они распространяются из активизировавшихся реликтовых очагов чумы.
Клиника болезни у человека или грызуна, в основном предопределяется паразитированием Y. pestis среди макрофагов, эволюционных потомков свободно живущих одноклеточных организмов. Возбудитель из места проникновения (например, места укуса чумной блохи) заносится в ближайшие лимфатические узлы, и они воспаляются. Из кровеносной системы в инфицированный лимфатический узел рекрутируются другие макрофаги, так формируется бубон, заполненный останками погибших макрофагов. Размножившиеся Y. pestis поступают из лимфоузлов в кровь (септицемия); выброс разрушающимися макрофагами огромного количества лимфокинов, нехарактерного для иммунных реакций, протекающих в варианте нормы и к которым адаптирован организм, дает картину шока и помрачения сознания больных. У людей отдельных генотипов развиваются кожные (карбункулы, петехии, геморрагии) и/или легочные поражения (вторичная легочная чума). Так как процесс разрушения макрофагов протекает быстро и возбудитель чумы поступает в кровь, где он доступен действию антибиотиков, то своевременно начатая антибиотикотерапия обычно дает эффект (в отличие антибиотикотерапии при бруцеллезе, возбудитель которого вступает в эндосимбиотические отношения с макрофагами). Поэтому сегодня смертность при бубонной чуме составляет не более 8 %, вместо 50–80% в эпоху «до антибиотиков».
Таким образом, признание сапронозного характера существования в природе возбудителей опасных для человека инфекций, когда исключается «сапрофитическая фаза» в их существовании, а сами микроорганизмы рассматриваются в качестве паразитов или эндосимбионтов простейших организмов, дает возможность объяснять многие феномены их эпидемиологии, не укладывающиеся в модель «грызун-блоха-грызун». Это одновременно и компромисс с устоявшимися взглядами, и толчок для мысли и экспериментов на новом методическом уровне. Он потребует от ученых с одной стороны, осознания того, что существуют природные очаги инфекционных болезней иные, чем изучаемые сегодня в рамках представлений о природно-очаговых болезнях (о чем и свидетельствуют исторические источники); с другой, необходимость применения новых определений для описания таких очагов и эпидемиологии поддерживающихся в них патогенных микроорганизмов. В качестве рабочих терминов для описания эпидемических процессов вне учения о природной очаговости чумы Д. К. Заболотного, оставляющего основную роль в поддержании Y.pestis в природе за грызунами, мы предложили ряд определений (Супотницкий М.В., Супотницкая Н.С., 2006). Ниже я привожу их в формулировках, применимых и для других сапронозов:
реликтовый очаг опасной инфекционной болезни — территория, неопределенно долго включающая природный очаг возбудителя опасной инфекционной болезни, о существовании на которой в прошлом вспышек данной болезни среди людей и животных известно из исторических источников. Термин — гипотеза. Должен применяться при историческом описании эпидемий и при постановке задачи на определение методами молекулярной диагностики границ природных очагов возбудителей опасных инфекций, поддерживающихся среди простейших и не проявляющих себя эпизоотиями или вспышками инфекционной болезни среди людей в настоящее время;
природный резервуар возбудителя опасной инфекционной болезни (первичный) — совокупность одноклеточных организмов — биологических хозяев возбудителя опасной инфекционной, без которых его существование в природе как биологического вида, невозможно;
природный очаг возбудителя опасной инфекционной болезни (первичный) — географический ландшафт, в почве которого методами молекулярной диагностики доказано присутствие этого возбудителя среди простейших, но не проявляющий себя эпизоотиями или вспышками инфекционной болезни среди людей в настоящее время (холодный очаг), и/или на его территории фиксируются эпизоотии и эпидемии данной инфекционной болезни (пульсирующий или активизировавшийся очаг);
усилители природного резервуара возбудителя опасной инфекционной болезни — биотические объекты (растения, грызуны, их эктопаразиты) не имеющие значения для поддержания в природе возбудителя данной болезни как биологического вида, но способные накапливать, размножать и доставлять его в организм человека;
активизировавшийся природный очаг возбудителя опасной инфекционной болезни — продолжающееся в течение исторически зафиксированного периода времени (до нескольких столетий) появление на территории природного очага возбудителя опасной инфекционной болезни, ее эпизоотий и эпидемий;
пульсация природного очага возбудителя опасной инфекционной болезни — процесс кратковременного (до нескольких лет), интенсивного и охватывающего обширные территории разрушения экосистем «простейшие–возбудитель опасной инфекционной болезни», проявившийся проникновением этого микроорганизма в популяции диких и домашних грызунов и их эктопаразитов.
Проникновение возбудителя чумы из первичного резервуара в популяции животных и человека, — катастрофическая случайность для всех — и для микроорганизма, так как он попадает в среду, являющуюся для него биологическим тупиком; и для его жертв (человека, животных и их эктопаразитов). Но в результате разрушения экосистемы «простейшие—Y. pestis», возбудитель чумы «отрывается» от своего первичного природного резервуара и при наличии благоприятных условий (особенности биологии грызунов и их эктопаразитов, увеличение их численности, восприимчивости и др.) формирует вторичные резервуары. Продолжительность существования такого резервуара может не превышать продолжительности инфекционного процесса у отдельного инфицированного животного. Однако, в тех случаях, когда благодаря эктопаразитам грызунов между ними формируются эпизоотические (эпидемические — когда речь идет о человеке) цепочки, микроорганизм — облигатный паразит простейших, получает возможность длительно поддерживаться в наземном резервуаре. Основную роль в его поддержании играют макрофаги/моноциты грызунов, эволюционные потомки простейших. Попав в организм человека или грызуна, микроорганизм использует те же механизмы специализации, которые позволяют ему поддерживаться среди почвенных одноклеточных организмов. В результате среди животных, обитающих на определенной территории, вспыхивают эпизоотии чумы, сами они становятся источником Y. pestis для людей, т. е. усилителями ее первичного природного резервуара, но вся их совокупность формирует вторичный природный очаг Y. pestis.
Вследствие меньшей устойчивости вторичные природные очаги чумы не могут существовать дольше, чем первичные. После «угасания» таких очагов остается огромная территория — первичный очаг, в котором микроорганизм продолжает поддерживаться среди одноклеточных организмов.
«Третья пандемия чумы»
Так называют активизацию очагов чумы, пришедшуюся на конец XIX столетия и совпавшую с открытием А. Иерсеном (Yersin, 1863–1943) возбудителя чумы — Y. pestis. Теперь в эпидемиологии чумы стало все ясно и понятно, тогда же и появилась эта схема «грызун-блоха-грызун». Появилась и еще одна «каноническая схема», представленная на рисунке ниже. Об нее то «грызуноблохисты» и «ломают» всякого рода сторонников «ненаучных» идей в чумологии. Но вот насколько научной является сама эта схема, давайте разберемся.
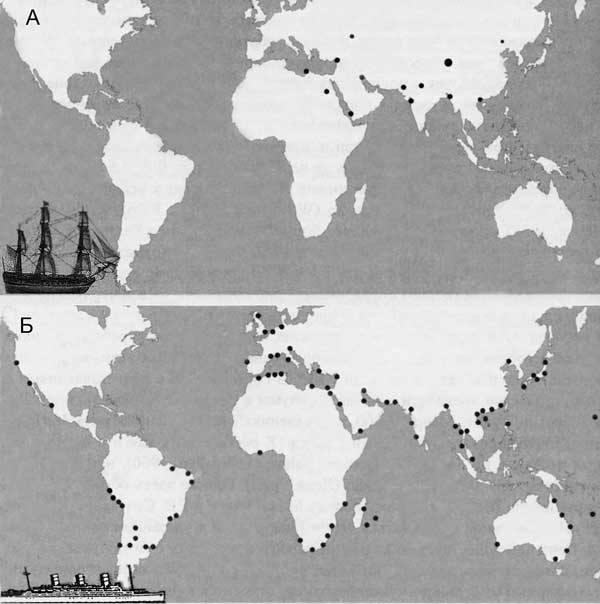
Схема распространения очагов чумы в мире по В. Н. Федорову, В. П. Козакевичу (1959). А — до начала третьей пандемии чумы с 1846 г. по 1894 г. Б — в начале третьей пандемии с 1894 г. по 1903 г. Любопытный образчик конъюнктурного искажения научной информацию в угоду господствующей в эпидемиологии чумы концепции. Проигнорированы предшествовавшие «третьей пандемии» вспышки чумы в горных долинах китайской провинции Юнъ-нан (о них было известно уже не менее 50 лет!), в Индии, Курдистане, Афганистане, Северной Персии, Астраханском крае, Месопотамии, в центральной Танзании и Уганде, в Маньчжурии, Забайкалье и в др. местностях. И куда только они делись с карты с началом «третьей пандемии»? Схема взята из источника 2006 г., где приведена как «образец научности» схемы «грызун-блоха-грызун».
Пандемии чумы начинаются за несколько десятилетий до своего «официального признания» в качестве эпидемической катастрофы — реликтовые очаги чумы разогреваются медленно. Так первая исторически зафиксированная пандемия чума (чума Юстиниана, 542–589) в восприятии современников сначала проявилась разрозненными случаями в Константинополе в 531 г. Историк церкви Евсевий Памфил (Eusebius, ок. 260–340) в своем труде «Церковная история» упоминает о чуме, как об известной (в отличие от оспы) на его время болезни.
Второй пандемии также предшествовали какие-то эпидемии, сопровождавшиеся повальной смертностью, и оставившие свидетельства в легендах и летописях именно как чума или мор. Культ Святого Рока как защитника от чумы, появился в Европе перед «черной смертью», сам святой умер в тюрьме в 1327 г., почти за 20 лет да начала «черной смерти». Традиция изображать «Гнев Господень» в виде стрел чумы, ниспосланных сверху, начинается еще раньше, как минимум, в XIII в. (Делюмо Ж., 1994). На территории России летописцы в течение двух столетий, предшествовавших «черной смерти», фиксировали «бысть мор» в городах, ставших потом ее основной жертвой (Смоленск, Псков, Новгород, Изборск и др.). «Мор» с завидной регулярностью начинался в конце лета (Рихтер В. М., 1814). С XIII в. «затлели» очаги чумы на Балканах, в Курдистане и Месопотамии.
В XIX в. угасая на Европейском континенте, очаги чумы разгораются южнее, в глубине территорий, занимаемых сегодня Индией и Китаем. Эти территории были интенсивно поражены чумой во время «черной смерти», но в XVII в. Индостан и Китай оставались относительно благополучными по чуме. В начале XIX в. чума в Индии вновь резко активизировалась. Бубонная и легочная чума появились в 1815 г. горной провинции Куче и на юго-востоке провинции Гузерат (штат Гуджарат — там же она появилась в 1994 г., и, разумеется, оказалась нераспознанной более месяца, так как о чуме в этих местах уже никто не помнил). В 1819 г. чума в Индии получила еще большее распространение. В 1836 г. она появилась в местности, отстоящей далеко от пораженных прежде, в Пали. В эти же годы началась активизация природных очагов чумы на юге Китая и в Индокитае. К 1834–1835 гг. относятся сведения о появившейся в китайском городе Нинпо (Нинбо — город на берегу Восточно-Китайского моря) эпидемии «чумоподобного характера». В 1850 г., когда бубонная чума выпала из поля зрения европейских ученых, и, даже, считалась ими «вымершей болезнью», легочная чума вновь вспыхнула на южных склонах Гималаев (в Гурвале и Камауне). В этот же год бубонная чума напомнила о себе в Кантоне (Китай). В 1858 г. обнаружилось «движение» чумы на север и запад. Она появилась в бубонной форме среди бедуинов на прибрежье Средиземного моря вблизи Триполи, где чума была известна еще во времена карфагенского владычества. Через 5 лет активизировались очаги Великого Евразийского чумного излома (1863–1879), с начала 1880-х гг. чума все чаще регистрируется в Северном Прикаспии, Поволжье, Монголии, Северном Китае и Забайкалье. Со средины XIX в. чума становится эндемичной на юге Китая. В 1864 г. вновь «чумоподобная эпидемия» в Нинпо. В 1866 г. чума обнаружена в столице китайской провинции Юнъ-нань (граничит с Вьетнамом и Бирмой, от самого южного ее участка до побережья моря не менее 400 км) — в городе Юнъ-нань-фу (до побережья не менее 1000 км). Во время гражданских войн в 1860–1870 гг. чума свирепствовала на юге Китая эндемически (!). В 1867 г. чума вспыхивает в Пакхое (город в провинции Гуанъ-дунъ на берегу Тонкинского залива), но никуда не «разносится кораблями и крысами». В 1871 г. чума «возвращается» в провинцию Юнъ-нань, продолжаются значительные вспышки чумы в Пакхое. С конца 70-х годов XIX в. обнаружилась сезонность в появлении чумы в южном Китае. Она стала аккуратно появляться в январе каждого года в провинции Квангси (Гуанъ-си) и в округах Лиенчоу и Лейчоу. Болезнь наблюдалась ежегодно с 1881 г. по 1884 г. в Пакхое, но кораблями она почему-то не распространяется. В 1889 г. была эпидемия в Люнгчоу (Лунчжоу), а в 1890 г. чума вспыхнула в Ву-чу (Бучу), на берегу между Пакхоем и Кантоном. В 1891 г. появилась чума в Као-чау, области смежной с Лиенчоу, где находится Пакхой. В 1892 г. вспыхнула эпидемия чумы в Ан-пу, городе, находящемся в 180 км к востоку от Пакхоя. В 1893 г. вновь появилась чума в Пакхое. Она также была в Юнъ-нане, преимущественно в городе Мингз (в южной части провинции). Чума свирепствовала в Люнгчоу и во многих глубинных городах провинции Квангси. В 1894 г. чума устраивает побоище в китайских портовых городах Кантоне, Гонконге и Амое, и только тогда (!) она попадает в поле зрения европейских ученых, и понимается ими как начавшаяся третья пандемия.
За первые 10 лет пандемии (1894–1903) чума отметилась в 87 (из нескольких тысяч!) портовых городов. Хотя в те же годы фиксировали эпидемии, которые не были связанны с морскими портами, все равно действовал определенный стереотип мышления: пандемия чумы началась в 1894 г. и была «портовой» (Ахшарумов Д. Д., 1900; Диаптроптов П. И., 1901; Wu Lien-Ten et al., 1936). Факты, свидетельствовавшие о том, что чума в XIX в не прекращалась, не принимались во внимание, так как они противоречили ставшей очень прогрессивной теории разноса чумы кораблями, кстати, известной еще со времен чумы в Марселе 589 г. (см. у Гезера Г., 1867). Следовательно, началом так называемой «третьей пандемии чумы» сегодня считается случайно взятая дата одной из эпидемий чумы в Кантоне, периодически вспыхивающих там с 1850-х гг., а схема «грызун-блоха-грызун», ставшая основой для объяснения распространения портовой чумы, существует только для того, что бы бесконечно вылавливать грызунов и вычесывать у них блох.
Изучение хронологии эпидемий по городам позволило выявить нам несколько пиков активности чумы в Европе во время ее второй пандемии — первый пик (1346–1382), второй (1440–1530), третий (1545–1683; максимум пандемии), четвертый (1710–1830) и пятый (1899–1931). Пики не равноценны. Четвертый даже не имел территориальных размахов первого. А пятый («третья пандемия чумы») дал в Европе всего несколько незначительных по смертности вспышек среди людей (Оппорто, 1899; Глазго, 1900; Неаполь, 1900; Константинополь, 1900, 1919–1929; Одесса, 1902, 1910; Париж, 1930; низовья реки Турия и долина реки Эбро на Пиренейском полуострове, 1931; и др.). Вспышки развились на местностях, уже известных как чумные в прошлом. Следовательно, эти пики чумы стали проявлением одного процесса активизации природных очагов чумы в Европе, имевшего свое начало (1346–1382), максимум (1545–1683) и угасание (1710–1830; 1899–1931).
Сопоставление же территориальных масштабов так называемой «третьей пандемии» с предыдущими пиками активности чумы второй пандемии, показывает, что она значительно уступает по этому показателю последнему, четвертому ее пику (1710–1830). Учитывая также и искусственность даты начала третьей пандемии, правильнее будет считать ее не самостоятельным явлением, а пятым, самым низким пиком второй пандемии чумы.
Клинические особенности эпидемий чумы в разные исторические периоды
Интерес для исследователя могут представлять различия в клинике чумы первой и второй пандемий.
По первой пандемии до нас дошло описание клиники чумы, оставленное историком ранней византийской эпохи, Прокопием. Если привести сделанное им разделение клинических форм болезни в соответствие с ее современной клинической классификацией Г. П. Руднева (1970), то они соответствуют бубонной и первично-септической формам чумы. Болезнь не передавалась между людьми и в тоже время было замечено, что заболеть чумой можно было без всяких контактов, например, заперевшись в своем доме (см. Гезер Г., 1867; Гиббон Э., по изд. 1997).
Описания клиники чумы у авторов-современников начала второй пандемии, различаются. Но почти все они (за исключением Дж. Боккаччо) отмечали такой клинический симптом, как кровохарканье. Он всегда ими рассматривался как признак скорой смерти больного, но, видимо, не был известен для чумы из прежней практики. Шольяк называл «черную смерть» чумой с кровохарканьем. С сильнейшими болями в груди больные чумой выхаркивали вещества, окрашенные кровью. Из их рта выходило болезненное зловоние, затем присоединялись сухость языка и глотки, неутолимая жажда, бессонница и мучительные, распространенные по всему телу ощущения. Преимущественно при этой форме болезни на коже человека выступали красные и черные пятна, различные по плотности и насыщенности цвета (карбункулы и петехии). Бубоны появлялись на руках, челюстях и в других частях тела. Трупы людей быстро чернели, отсюда и пошло название «черная смерть» (Гезер Г., 1867).
Описание Шольяка и других врачей того времени, собранные Г. Гезером (1867), свидетельствуют о том, что во время «черной смерти» типичные бубонные случаи болезни сопровождались легочными осложнениями, что по клинической классификацией Г.П. Руднева (1970) соответствует вторично-легочной чуме. Так же как и в первую пандемию, врачами были описаны бубонная и первично-септическая формы чумы, но легочные осложнения видимо были для них клинической новинкой. Если подойти к рассмотрению причин появления легочной чумы с точки зрения современных авторов, то ничего необычного в таком течении эпидемии нет. Ими обычно приводится процент случаев, сопровождавшихся осложнениями со стороны легких (см., например, у И. В. Домарадского, 1998).
Однако свидетельства очевидцев массовых эпидемий чумы XVI–XIX столетий показывают, что переход бубонной чумы во вторично-легочную не является закономерным и не зависит от количества людей, заболевших бубонной чумой. А. Гирш (1853), занимавшийся изучением клиники чумы по первоисточникам, заметил, что, начиная с XVI в., легочные осложнения вообще исчезли из описаний эпидемий чумы. Какие бы масштабы эпидемия не принимала (Лондон, 1665 г., Москва, 1771–1772 гг. — почти по 100 тыс. погибших от чумы в каждой эпидемии), болезнь развивалась исключительно в бубонной форме (детальное описание на русском языке чумы в Москве см. в работах А. Ф. Шафонского и Д. Самойловича; в Лондоне, в работе Д. Дефо). Легочные осложнения для нее считались «нехарактерными». На Руси последние эпидемии легочной чумы зафиксирован в 1424–1425 гг. в Твери, Москве, Новгороде, Пскове, Карелии, а также в «Литве и Немцах» (Рихтер В. М., 1814). О легочной чуме постепенно забыли, и когда она стала появляться отдельными вспышками в XIX в. среди изолированных групп населения, то среди ученых начались дискуссии о самой возможности такой чумы. И только обращение к источникам времен «черной смерти» позволило установить, что речь идет об одной и той же болезни, но протекающей в разных клинических формах. Вполне логично предположить, что люди, жившие в Европе в VI в., значительно отличались по генотипам, от живших там же в XIV в., и что именно люди отдельных генотипов дают легочные осложнения при бубонной чуме. Во время чумы «черной смерти» и при последующих вспышках болезни, они были элиминированы из этносов, живущих на территориях реликтовых очагов чумы, поэтому вспышки легочной чумы там прекратились на сотни лет.
Для иллюстрации данного положения, обратимся к исследованию Г. Н. Минха (1898) динамики развития чумы 1878 г. в станице Ветлянская (Астраханская область). По клиническим проявлениям эта эпидемия делится им на два периода: 1) с преобладанием бубонных форм болезни (с 28 сентября); 2) с преобладанием безбубонных и, в основном, легочных форм (с 27 ноября). Второй период эпидемии начался в многочисленной семье казака Осипа Белова. Эпидемия проявила себя сразу массой скоротечных смертельных заболеваний с почти полным отсутствием бубонов у заболевших чумой. Но Минх констатировал, что близкое соседство даже с наиболее пострадавшими от чумы семействами, не имело никакого влияния на распространение болезни по станице. Ему также удалось установить, что основной принцип ее «движения» — не соседские или внутридомовые отношения между заболевшими, а родственные. В его исследовании приведены десятки родословных местных жителей. Минх не знал таких терминов как генотип или геном, поэтому он сделал вывод, что «легочная чума распространяется по родословным». Следовательно, второй период ветлянской чумы начался со случайного вовлечения в эпидемический процесс особых человеческих генотипов, у которых инфекционный процесс проявился вторично-легочной формой болезни. После достижения «критической массы» из вторично-легочных случаев болезни, чума сформировала самостоятельные эпидемические цепочки, состоящие уже из первично-легочных случаев болезни. К удивлению врачей эпидемия в конце декабря внезапно прекратилась сама, как бы «поразив все намеченные объекты».
Но при ближайшем рассмотрении некоторых вспышек чумы могут появиться и другие гипотезы, уже касающиеся экологии самой Y. pestis. Интересно свидетельство Гюи Шольяка об эпидемии в Авиньоне в 1348 г. Он также как и Минх, разделил ее на два клинически различных периода. В первом, продолжавшемся два месяца, кроме лихорадки (в понимании врачей того времени) основным симптомом становилось кровохарканье. Продолжительность жизни больного не превышала 3-х суток. Во втором периоде эпидемии болезнь протекала преимущественно в бубонной форме, больной погибал в течение 5 суток (см. Гезер Г., 1867). Можно поставить под сомнение описание Шольяка, но сходным образом развивалась чума в 1897 г. в горном кишлаке Анзоб, расположенном в отрогах Гиссарского хребта (сегодня Таджикистан), описание которой оставил Я. М. Финкельштейн (1906). Следовательно, такое течение эпидемий чумы — одна из возможных закономерностей в ее развитии. Но по своей сути они представляют собой как бы «зеркальной отражение» чумы в станице Ветлянская. Чума начинается не с бубонных форм, затем переходящих сначала во вторично-легочные, а потом в первично-легочные (Ветлянка), а наоборот, вспышка чумы начинается с легочных форм болезни, заканчивается бубонными (Авиньон, Анзоб). Значит и механизмы развития чумы в Авиньоне и Анзобе могут отличаться от механизмов ветлянской чумы.
По чуме в Авиньоне и Анзобе можно дать два объяснения. Первое, наиболее вероятное, это преобладающее вовлечение в эпидемию людей тех генотипов, у которых бубонная форма болезни быстро переходит во вторично-легочную, и их последующая элиминация. Такие люди изначально представляли собой значительную прослойку среди населения Авиньона или кишлака Анзоб, а не были как в станице Ветлянская, лишь «вкраплением» в общей популяции. Второе объяснение, более спорное, а поэтому и более интересное — вовлечение в эпидемию штаммов возбудителя чумы, имеющих большую тропность к макрофагам легочной ткани. Селекция таких штаммов Y. pestis могла быть результатом их паразитирования в простейших, имеющих рецепторы, сходные с теми, что представлены на поверхности легочных макрофагов. Возможность получения пневмотропных штаммов возбудителя чумы подтверждена экспериментально в начале 1930-х гг. профессором А. М. Скородумовым при изучении штамма, выделенного в 1927 г. во время эпидемии легочной чумы в Монголии. Подкожное и внутрибрюшинное заражение таким штаммом морской свинки вызывало у нее пневмонию (Шунаев В. В., 1934).
***
Последние несколько лет правительство выделяет значительно больше денег на развитие науки, чем это имело место в 1990-х гг. В противочумной системе идет ремонт зданий, закупается новое научное оборудования. Но в разговорах с некоторыми сотрудниками я слышал обеспокоенность старением научных коллективов и нежеланием молодежи идти в науку. Но под какие идеи мы будем привлекать молодежь к исследованию чумы? Современная чумология строится на достижениях, полученных еще в доантибиотическую эпоху. Наличие неприкасаемых концепций типа рассмотренных в этой статье, привело к самоизоляции чумологии от многих современных направлений в исследовании экологии микроорганизмов. Нельзя сказать, что такое положение в чумологии не замечено известными учеными. Еще в 1998 г. И. В. Домарадский констатировал кризис в исследованиях чумы. Чумология впала в частности, и в их бесконечные уточнения, а в результате вся имеющаяся в ее распоряжении теоретическая база оказалась не способной объяснить поддержание Y. pestis на отдельной территории без эпизоотий грызунов всего лишь в течение периода, чуть более 10 лет (см. у И. В. Домарадского, 1998). Исторические источники дают нам представления о чуме, как о явлении значительно более сложном, чем отдельная чумная эпизоотия среди грызунов. Однако у исследователей нет мотивации делать научные открытия. Так в период «борьбы с биотерроризмом» конъюнктурные объяснения причин второй пандемии чумы в Европе якобы применением биологического оружия неким татарским ханом, всех устраивали, так как они открывали новые возможности по финансированию. Тем не менее, конъюнктурные объяснения не могут заменить знаний реальных механизмов возникновения масштабных эпидемий чумы в Европе в течение почти 500 лет. Никто не пытался объяснить связь в появлении пандемий оспы, проказы и чумы, разное клиническое течение чумы в разные исторические эпохи и еще ряд феноменов, присутствующих в исторических описаниях эпидемий чумы. А они существуют в объективной реальности. Те объяснения, которые мы им дали в монографии «Очерки истории чумы» (2006), разумеется, несовершенны, но давайте их поищем вместе. Выход на уровень новых гипотез даст мощную мотивацию для научной деятельности молодым сотрудникам.
Январь 2009 г.
Литература
Анисимов А. П. Факторы Yersinia pestis, обеспечивающие циркуляцию и сохранение возбудителя чумы в экосистемах природных сообществ. Сообщения 1 и 2 // Молекулярная генетика, микробиология и вирусология. — 2002. — № 3. — С. 3–23; № 4. С. 3–11.
Архангельский Г. Ф. Амбулантные формы чумы (pestis ambulans) и ее значение в эпидемиологии // Сборник сочинений по судебной медицине, судебной психиатрии, медицинской полиции, общественной гигиене, эпидемиологии, медицинской географии и медицинской статистике. — 1879. — Т. 1. — С. 132–193.
Ахшарумов Д. Д. Чума последних годов XIX столетия. — Полтава, 1900.
Белиловский В. А., Гамалея Н. Ф., Бурда М. К. Чума в Одессе. Т.1., Т.2. — Одесса, 1904.
Боккаччо Дж. Декамерон. — Кишинев, 1982.
Борисенков Е. П., Пасецкий В. М. Тысячелетняя летопись необычайных явлений природы. — М, 1988.
Бразоль Л. Е. Дженнеризм и Пастеризм, критический очерк оснований оспопрививания. — Харьков, 1875.
Бухарин О. В. Персистенция патогенных бактерий. — М., 1999.
Бухарин О. В., Литвин В. Ю. Патогенные бактерии в природных экосистемах. — Екатеринбург, 1997.
Васильев К. Г., Сегал А. Е. История эпидемий в России. — М., 1960.
Веревкин И. История оспы в России. — СПб., 1867.
Вогралик Г. Ф. Учение об эпидемических заболеваниях. — Томск, 1935.
Гезер Г. История повальных болезней. — СПб., 1867.
Гиббон Э. Закат и падение Римской империи. — М., 1997.
Гирш А. Индийская чума // Военно-медицинский журнал. — 1853. — Часть LXII (III). — С. 29–48.
Дербек Ф. И. История чумных эпидемий в России. — СПб., 1905.
Делюмо Ж. Ужасы на западе. — М., 1994.
Дефо Д. Дневник чумного года. — М., 1997.
Диаптроптов П. И. Обзор заболеваний чумой на побережье Средиземного моря и в портовых городах Европы в 1900 и в начале 1901 гг. // Русский архив патол., клин., мед. и бактериол. — 1901. — Т. 12. — С. 77–94.
Домарадский И. В. Чума. — М., 1998.
Жданов В. М. Эволюция вирусов // Природа. — 1988. — № 5, С. 4–14.
Киттеридж Дж. Л. Колдовство в Средние века. — СПб., 2005.
Козлов М. П., Султанов Г. В. Эпидемические проявления чумы в прошлом и настоящем. — Махачкала, 1993.
Купер Э. Сравнительная иммунология. — М., 1980.
Литвин В. Ю., Пушкарева В. И. Факторы патогенности бактерий: функции в окружающей среда // Журн. микробиол. — 1994. — приложение. — С. 83–87.
Минх Г. Н. Чума в России (Ветлянская эпидемия 1878–1879 гг.). — Киев, 1898.
Никульшин С. В., Онацкая Т. Т., Луканина Л. М. Изучение ассоциаций почвенных амеб H. rhisodes с бактериями - возбудителями чумы и псевдотуберкулеза в эксперименте // Журн. микробиол. — 1993. — № 9–10. — С. 2–5.
Онищенко Г. Г., Шапошников А. А., Субботин В. Г. с соавт. Противодействие биологическому терроризму. — М., 2003.
Пушкарева В. И. Патогенные бактерии в почвенных и водных сообществах (экспериментально — экологическое исследование). Автореф. дисс... д-ра биол. наук. — М., 1994.
Рихтер А. История медицины в России. — М., 1814.
Самойлович Д. Рассуждения о чуме, производившей в 1771 г. опустошения в Российской империи и особенно в столичном городе Москве // Избранные произведения. — М., 1952. — Т. 2. — С. 8–278.
Сунцов В. В., Сунцова Н. И. Чума. Происхождение и эволюция эпизоотической системы. — М., 2006.
Супотницкий М. В., Супотницкая Н. С. Очерки истории чумы. — М., 2006.
Супотницкий М. В. Эволюционная патология. — М., 2009.
Трахтенберг Дж. Дьявол и евреи. Средневековые представления о евреях и их связь с современным антисемитизмом. — М., 1998.
Федоров В. Н., Козакевич В. П. Современное распространение чумы в зарубежных странах // Природная очаговость и эпидемиология особо опасных инфекционных болезней. — Саратов, 1959. — С. 18–39.
Финкельштейн Я. М. Еще раз о чуме в Анзобе // Военно медицинский журнал. — 1906. — Т. 217, № 9. — С. 27–57.
Черкасский Б. Л. Сибирская язва как биологическое оружие. — М., 2002.
Шафонский А. Ф. Описание моровой язвы, бывшей в столичном городе Москве с 1770 по 1772 год, с приложением всех для прекращения оной тогда установленных учреждений. — М., 1775.
Шунаев В. В. // Сб. работ противочумных организаций Восточносибирского края за 1932–1933 гг. / Под ред. А. М. Скородумова, Москва, Иркутск, 1934.
Щепотьев Н. К. Чумные и холерные эпидемии в Астраханской губернии. — Казань, 1884.
Щепотьев Н. К. Чумные эпидемии в России. — СПб., 1897.
Cavanaugh D. C., Randall R. The role of multiplication of Pasteurella pestis in mononuclear phagocytes in the pathogenesis of fleaborne plague // J. Immunol. — 1959. — Vol. 85. — P. 348–363.
Cirillo J. D., Tompkins L. S., Falkow S. Growth of Legionella pneumophila in Acanthamoeba castellanii enhances invasion // Infect. Immun. — 1994. — Vol. 62. — P. 3254–3261.
Essig A., Heinemann M., Simnacher U. et al. Infection of Acanthamoeba castellanii by Chlamydia pneumoniae // Appl. Environ. Microbiol. — 1997. — Vol. 63. — P. 1396–1399.
Harb O., Lian-Yong Gao, KwaikY. A. From protozoa to mammalian cells: a new paradigm in the life cycle of intracellular bacterial pathogens // Environment. Microbiol. — 2000. — Vol. 2, № 3. — P. 251–265.
Marrie T. J., Raoult D., La Scola B. et al. Legionella-like and Other Amoebal Pathogens as Agents of Community-Acquired Pneumonia // Emerg. Infec. dis. — 2001. — Vol. 7, № 6. — P. 1026–1029.
Grabenstein J., Fukuto H., Palmer L. et al. Characterization of phagosome trafficking and identification of PhoP-regulated genes important for survival of Yersinia pestis in macrophages // Infection and Immunity. — 2006. — Vol. 74, № 7. — P. 3727-3741.
Greub G., Raoult D. Microorganisms resistant to free-living Amoebae // Clin. Microbiol. Rev. — 2004, — Vol. 17, No. 2. — P. 413–433.
La Scola B., Audic S., Robertet C. et al. A giant virus in amoebae // Science. — 2003. — Vol.299, № 5615. — P. 2033.
La Scola B., Raoult D. Survival of Coxiella burnetii within free-living amoeba Acanthamoeba castellanii // Clin. Microbiol. Infect. — 2001. — Vol. 7, № 2. — P. 75–79.
Wheelis M. Biological Warfare at the 1346 Siege of Caffa // Emerg. Infec. dis. — 2002. — Vol. 8, № 9. — P. 971–975.
Wu Lien-Ten U. A., Chun J. W., Pollitzen R. et al. Plague. A manual for medical and public health workers. — Shanghai, 1936.
Библиографическое описание:
Супотницкий М. В. Историческая информация о вспышках чумы как источник идей для будущих научных открытий в чумологии // Энвайронментальная эпидемиология и медицинская география. Ежегодник. Под ред. д. г. н. Д. В. Николаенко, Киев, 2011. — С. 298-329.

Михаил Васильевич Супотницкий
Российский микробиолог, полковник медицинской службы запаса, изобретатель, автор книг и статей по истории эпидемий чумы и других особо опасных инфекций, истории разработки и применения химического и биологического оружия. Заместитель главного редактора научно-практического журнала «Вестник войск РХБ защиты» Министерства обороны РФ.